|
Special Issue:
SPECIAL TOPIC — Modeling and simulations for the structures and functions of proteins and nucleic acids
|
| TOPICAL REVIEW—Modeling and simulations for the structures and functions of proteins and nucleic acids |
Prev
Next
|
|
|
Twisting mode of supercoil leucine-rich domain mediates peptide sensing in FLS2–flg22–BAK1 complex |
| Zhi-Chao Liu(刘志超)1,3, Qin Liu(刘琴)4, Chan-You Chen(陈禅友)4, Chen Zeng(曾辰)3, Peng Ran(冉鹏)1,3, Yun-Jie Zhao(赵蕴杰)2,†, and Lei Pan(潘磊)4,‡ |
1 School of Biological Information, Chongqing University of Posts and Telecommunications, Chongqing 400065, China
2 Institute of Biophysics and Department of Physics, Central China Normal University, Wuhan 430079, China
3 Department of Physics, The George Washington University, Washington, DC, 20052, USA
4 School of Life Sciences, Jianghan University, Wuhan 430056, China |
|
|
|
|
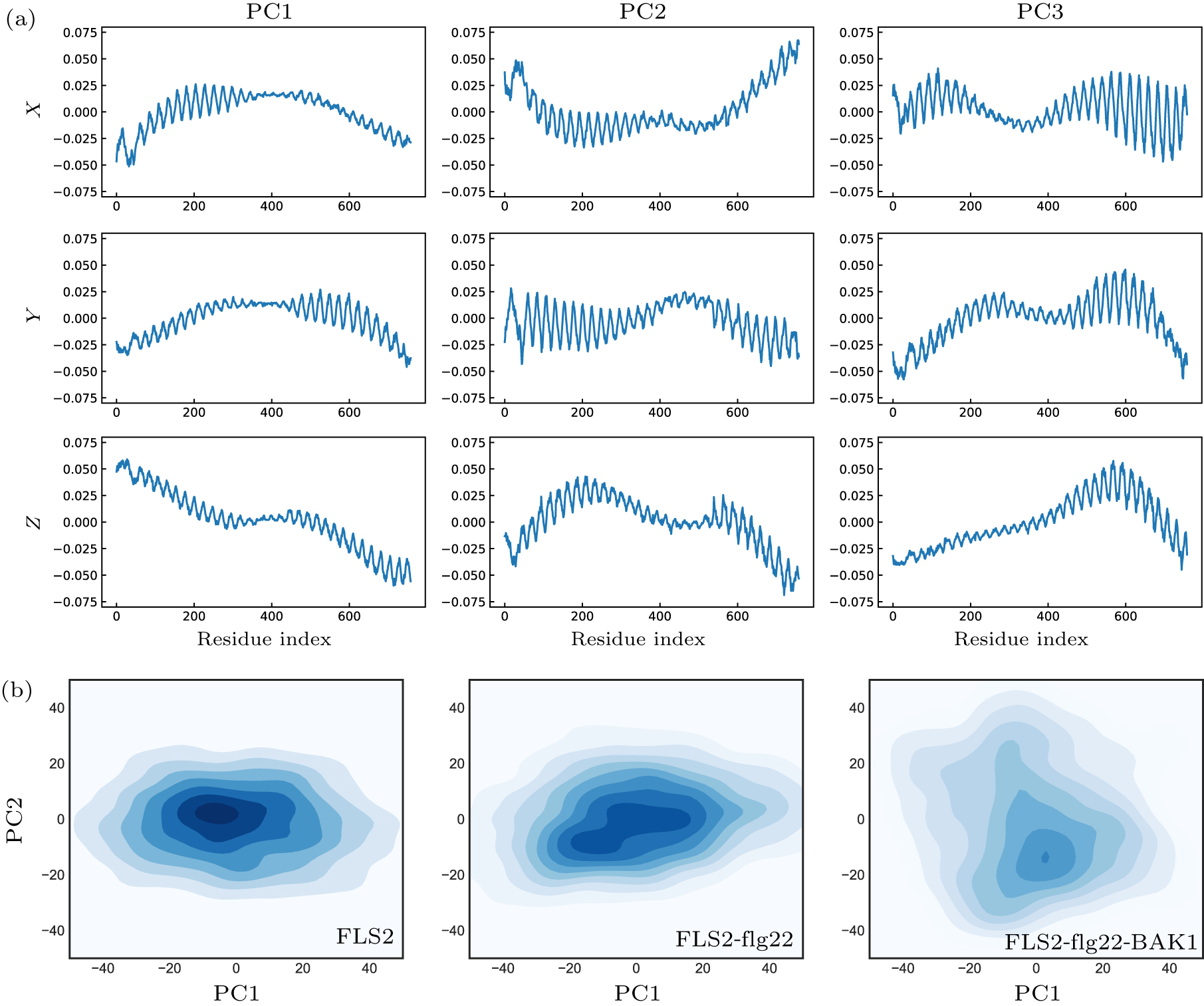
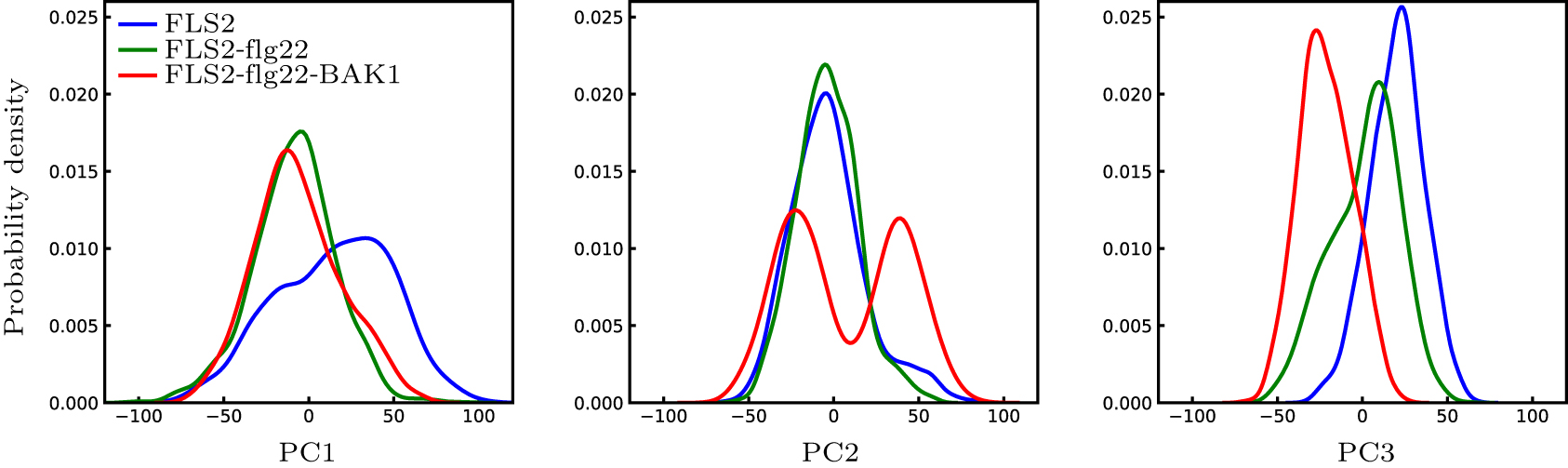
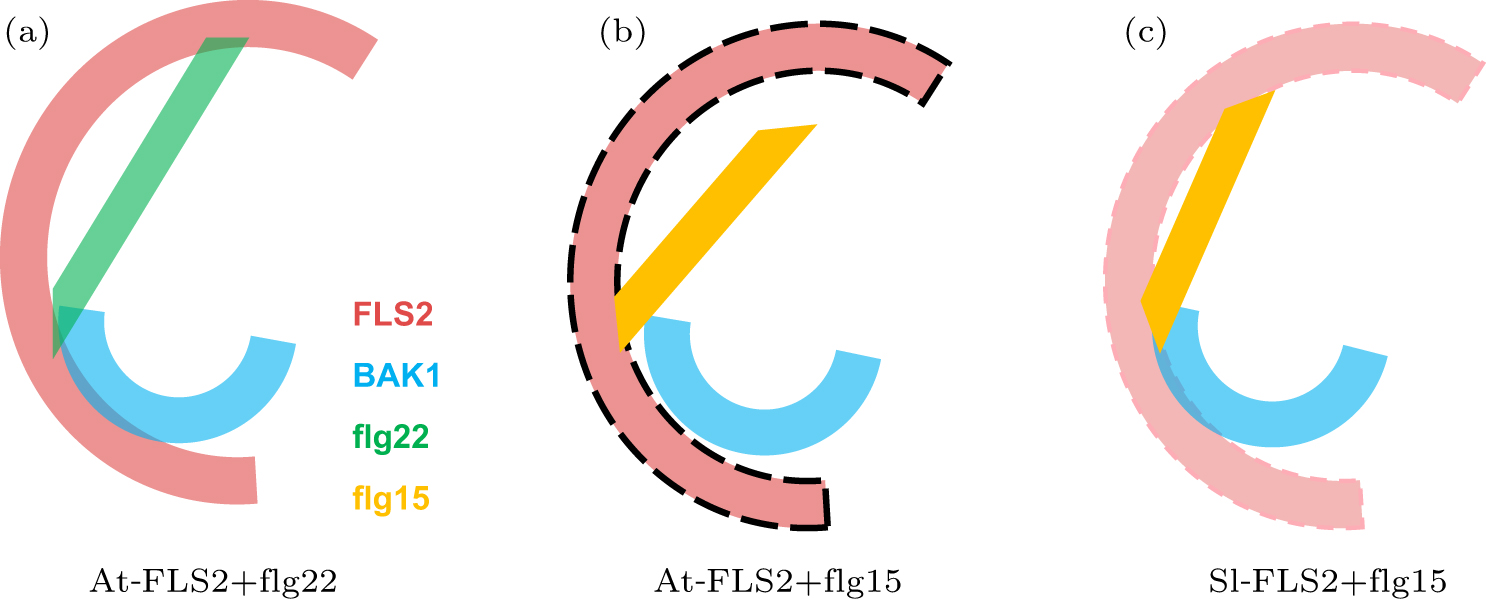
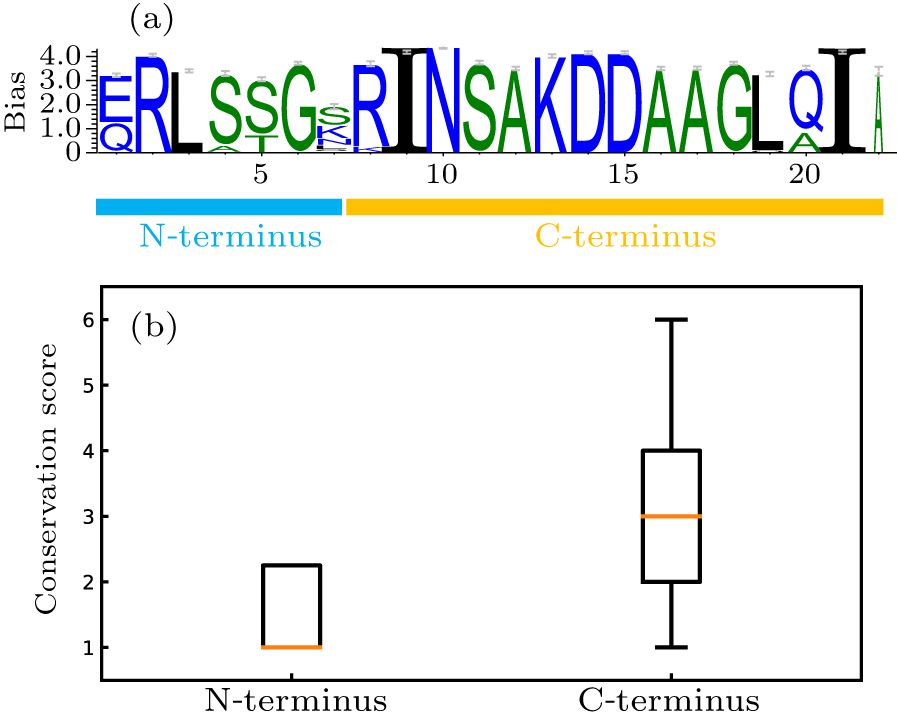
Abstract Plants and animals recognize microbial invaders by detecting pathogen-associated molecular patterns (PAMPs) through pattern-recognition receptors (PRRs). This recognition plays a crucial role in plant immunity. The newly discovered protein in plants that responds to bacterial flagellin, i.e., flagellin-sensitive 2 (FLS2), is ubiquitously expressed and present in many plants. The association of FLS2 and BAK1, facilitated by a highly conserved epitope flg22 of flagellin, triggers such downstream immune responses as activated MAPK pathway and elevated reactive oxygen species (ROS) for bacterial defense and plant immunity. Here we study the intrinsic dynamics and conformational change of FLS2 upon the formation of the FLS2–flg22–BAK1 complex. The top intrinsic normal modes and principal structural fluctuation components are very similar, showing two bending modes and one twisting mode. The twisting mode alone, however, accounts for most of the conformational change of FLS2 induced by binding with flg22 and BAK1. This study indicates that flg22 binding suppresses FLS2 conformational fluctuation, especially on the twisting motion, thus facilitating FLS2–BAK1 interaction. A detailed analysis of this sensing mechanism may aid better design on both PRR and peptide mimetics for plant immunity.
|
Received: 23 June 2020
Revised: 07 August 2020
Accepted manuscript online: 13 August 2020
|
|
PACS:
|
87.14.gn
|
(RNA)
|
| |
87.15.K-
|
(Molecular interactions; membrane-protein interactions)
|
| |
87.10.Ca
|
(Analytical theories)
|
| |
87.15.A-
|
(Theory, modeling, and computer simulation)
|
|
|
Corresponding Authors:
†Corresponding author. E-mail: yjzhaowh@mail.ccnu.edu.cn ‡Corresponding author. E-mail: leipan@jhun.edu.cn
|
| About author: †Corresponding author. E-mail: yjzhaowh@mail.ccnu.edu.cn ‡Corresponding author. E-mail: leipan@jhun.edu.cn * Project supported by the National Natural Science Foundation of China (Grant No. 11704140), self-determined research funds of CCNU from the Colleges’ Basic Research and Operation of MOE (Grant No. CCNU20TS004) (Y. Z.), and the China Scholarship Council Fund (Grant No. 201708420039) (L. P.). |
Cite this article:
Zhi-Chao Liu(刘志超), Qin Liu(刘琴), Chan-You Chen(陈禅友), Chen Zeng(曾辰), Peng Ran(冉鹏), Yun-Jie Zhao(赵蕴杰)†, and Lei Pan(潘磊)‡ Twisting mode of supercoil leucine-rich domain mediates peptide sensing in FLS2–flg22–BAK1 complex 2020 Chin. Phys. B 29 108709
|
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
Tintor N, Ross A, Kanehara K, Yamada K, Fan L, Kemmerling B, Nurnberger T, Tsuda K, Saijo Y 2013 Proc. Natl. Acad. Sci. USA 110 6211 DOI: 10.1073/pnas.1216780110 |
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
Chinchilla D, Zipfel C, Robatzek S, Kemmerling B, Nurnberger T, Jones J D, Felix G, Boller T 2007 Nature 448 497 DOI: 10.1038/nature05999 |
| [18] |
Somssich M, Ma Q, Weidtkamp-Peters S, Stahl Y, Felekyan S, Bleckmann A, Seidel C A, Simon R 2015 Sci. Signal 8 ra76 DOI: 10.1126/scisignal.aab0598 |
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
Zhao Y, Jian Y, Liu Z, Liu H, Liu Q, Chen C, Li Z, Wang L, Huang H H, Zeng C 2017 Sci. Rep. 7 2876 DOI: 10.1038/s41598-017-03003-5 |
| [28] |
Chen H, Zhao Y J, Li H T, Zhang D Y, Huang Y Z, Shen Q, Van Duyne R, Kashanchi F, Zeng C, Liu S Y 2014 Plos One 9 DOI: 10.1371/journal.pone.0109154 |
| [29] |
|
| [30] |
|
| [31] |
Brunette T J, Parmeggiani F, Huang P S, Bhabha G, Ekiert D C, Tsutakawa S E, Hura G L, Tainer J A, Baker D 2015 Nature 528 580 DOI: 10.1038/nature16162 |
| [32] |
Enkhbayar P, Kamiya M, Osaki M, Matsumoto T, Matsushima N 2004 Proteins 54 394 DOI: 10.1002/prot.10605 |
| [33] |
|
| [34] |
Ramisch S, Weininger U, Martinsson J, Akke M, Andre I 2014 Proc. Natl. Acad Sci. USA 111 17875 DOI: 10.1073/pnas.1413638111 |
| [35] |
Doyle L, Hallinan J, Bolduc J, Parmeggiani F, Baker D, Stoddard B L, Bradley P 2015 Nature 528 585 DOI: 10.1038/nature16191 |
| [36] |
|
| [37] |
Van Der Spoel D, Lindahl E, Hess B, Groenhof G, Mark A E, Berendsen H J 2005 J. Comput. Chem. 26 1701 DOI: 10.1002/(ISSN)1096-987X |
| [38] |
Duan Y, Wu C, Chowdhury S, Lee M C, Xiong G, Zhang W, Yang R, Cieplak P, Luo R, Lee T, Caldwell J, Wang J, Kollman P 2003 J. Comput. Chem. 24 1999 DOI: 10.1002/(ISSN)1096-987X |
| [39] |
Sethi A, Eargle J, Black A A, Luthey-Schulten Z 2009 Proc. Natl. Acad. Sci. USA 106 6620 DOI: 10.1073/pnas.0810961106 |
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
Ashkenazy H, Erez E, Martz E, Pupko T, Ben-Tal N 2010 Nucleic Acids Res. 38 W529 DOI: 10.1093/nar/gkq399 |
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
Mueller K, Bittel P, Chinchilla D, Jehle A K, Albert M, Boller T, Felix G 2012 Plant Cell 24 2213 DOI: 10.1105/tpc.112.096073 |
| [49] |
|
| No Suggested Reading articles found! |
|
|
Viewed |
|
|
|
Full text
|
|
|
|
|
Abstract
|
|
|
|
|
Cited |
|
|
|
|
Altmetric
|
|
blogs
Facebook pages
Wikipedia page
Google+ users
|
Online attention
Altmetric calculates a score based on the online attention an article receives. Each coloured thread in the circle represents a different type of online attention. The number in the centre is the Altmetric score. Social media and mainstream news media are the main sources that calculate the score. Reference managers such as Mendeley are also tracked but do not contribute to the score. Older articles often score higher because they have had more time to get noticed. To account for this, Altmetric has included the context data for other articles of a similar age.
View more on Altmetrics
|
|
|