








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Theoretical study of overstretching DNA–RNA hybrid duplex
Cite this Article
Yang Dong-Ni, Zhong Zhen-Sheng, Liu Wen-Zhao, Rujiralai Thitima, Ma Jie. Theoretical study of overstretching DNA–RNA hybrid duplex. Chinese Physics B, 2019, 28(6): 068701 
Permissions
Theoretical study of overstretching DNA–RNA hybrid duplex
† Corresponding author. E-mail:
Abstract
DNA–RNA hybrid (DRH) plays important roles in many biological processes. Here, we use a thermodynamic theory to analyze the free energy and unpeeling properties of the overstretching transition for the DRH molecule and compare the results with double-helix DNA. We report that the RNA strand of DRH is easier to get unpeeled than the DNA strand while the difficulty in unpeeling the double helix DNA lies in between. We also investigate the sequence effect, such as GC content and purine content, on the properties of unpeeling the DRH. Further, to study the temperature effect, the force-temperature phase diagram of DRH and DNA are calculated and compared. Finally, using a kinetic model, we calculate the force–extension curves in the DRH stretching and relaxation process under different pulling rates and temperatures. Our results show that both pulling rate and temperature have important influences on the stretching and relaxation kinetics of unpeeling the DRH. Putting all these results together, our work provides a comprehensive view of both the thermodynamics and kinetics in DRH overstretching.
1. Introduction
DNA–RNA hybrid (DRH) duplex is an important element in various biological processes such as transcription, reverse transcription[1] and gene editing.[2] It also plays essential roles in replication initiation,[3] gene regulation,[4] chromosome segregation,[5] DNA repair,[6] DNA methylation,[7] telomere elongation,[8] etc. In addition, the DRH duplex is also used in some biotechnologies such as antisense therapy,[9] and is related to human diseases including neurological disorders.[10]
Despite its significance, the mechanical properties of DRH have not been well characterized until very recently. Using magnetic tweezers, Zhang et al.[11] have measured the elasticity and the overstretching transitions of a single DRH molecule. Although the thermal stability of DRH[12–15] as well as its structure[16–19] (which lies between A-form and B-form) has been reported previously, single molecule stretching experiments can measure the force–extension relationship of DRH and thus provide a more direct and quantitative way to characterize its mechanical properties. Yet, to better understand the single molecule experimental data and, furthermore, to predict some mechanical properties of DRH which may not have even been experimentally explored, a theoretical investigation is clearly valuable and demanding.
In this paper, using both thermodynamic and kinetic model, we investigate the competition among different processes, e.g., S-form transition, unpeeling and bubbling which may occur in DRH overstretching as shown in Fig.
 | Fig. 1. Illustration of possible forms of DRH in its overstretching experiment: (a) double-helix DRH, (b) S-DRH, (c) unpeeled DRH (DNA-strand-stressed), (d) unpeeled DRH (RNA-strand-stressed), and (e) bubbled DRH. |
2. Theoretical model
We refer to a theoretical model previously proposed by Cocco et al.[20] to study the mechanical overstretching of dsDNA and extend it to describe the overstretching of DRH with the newly measured elastic parameters. For tension-driven experiments, the free energy per base pair at a given force can be described by the integration of extension from force–extension data as follows:
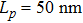
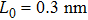
Zhang et al. have also shown that in the force region of 57–72 pN, DRH molecule underwent the structural transition and transformed from double helix form to S form, which, similar to the S form of dsDNA or double-stranded RNA (dsRNA),[22] is extended form of DRH with base-pairing retained as shown in Fig.
To describe the force–extension response at the double-helix to S-form transition stage and later the S-form stretching stage, we make an approximation that the extension of DRH is linearly dependent on force in these two scenarios but with different slopes, S1 and S2. The extension of DRH as a function of force at the double-helix to S-form transition stage thus can be written as
In addition, to make a comparison of relative free energy between single-stranded form and double helix form, we must consider the base-pairing free energy g0, which has a positive value here to denote the energy cost for opening one base-pa ir. It is noted that g0 could have been estimated from DRH unzipping experiments, which have not been reported yet. Alternatively, we refer to the base-pairing-stacking energy from Sugimoto et al.,[24] under the nearest-neighboring approximation as shown in Table 

| Table 1.
Base-pairing-stacking free energy of hybrid duplex from Sugimoto et al.[24] for 25 °C, 1-M NaCl. . |
Next, we analyze the stabilities of different forms of DRH by calculating the relative free energy between them. In the DRH stretching experiment, when unpeeling happens, if the RNA strand gets unpeeled and the DNA strand is left with tension, as shown in Fig.


To compare the stability of unpeeled DRH with that of DRH in S-form, the free energy of single-stranded form DNA or RNA relative to that of S-form DRH can be calculated as follows:


To consider whether tension-driven bubbling (Fig.
With Eqs. (
Further, using a kinetic model, we can simulate and predict the force–extension response of DRH in its stretching and relaxation experiments. As tension increases, unpeeling can occur from the nicks on the strands or from the open ends of the strands as shown in Figs.
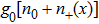


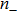

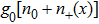


In addition, for simplicity, here we use wds to denote any double stranded form of DRH and therefore, it depends on the force as expressed below.

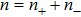
3. Results and discussion
In this section, we will present some representative results about the calculated free energy of DRH in its stretching experiment based on the model mentioned above. We will then discuss the overstretching thermodynamics of DRH and also make a comparison with double-helix DNA (i.e., B-DNA).
We first calculate the force-dependent free energy of double-helix DRH and DNA as a function of stretching force using Eqs. (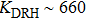
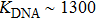

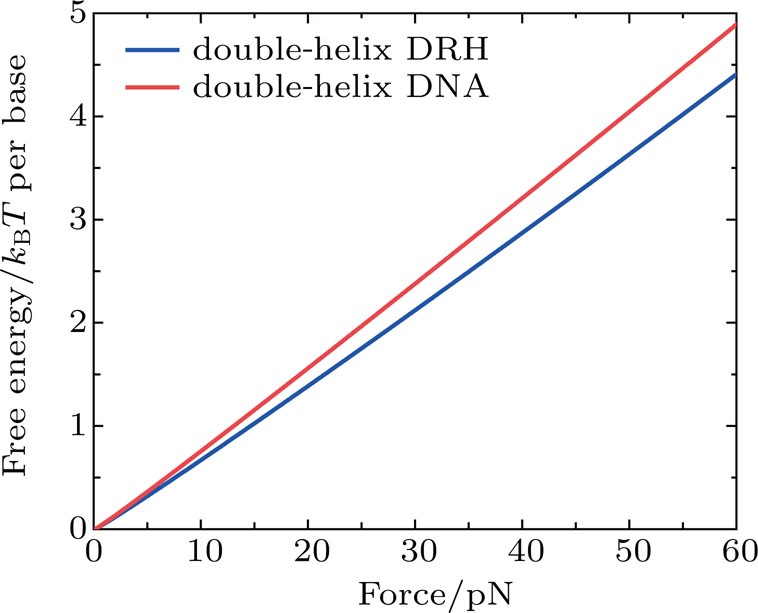 | Fig. 2. Free energy per base of double-helix DRH (wdhDRH) and double-helix DNA (wdhDNA) as a function of force. |
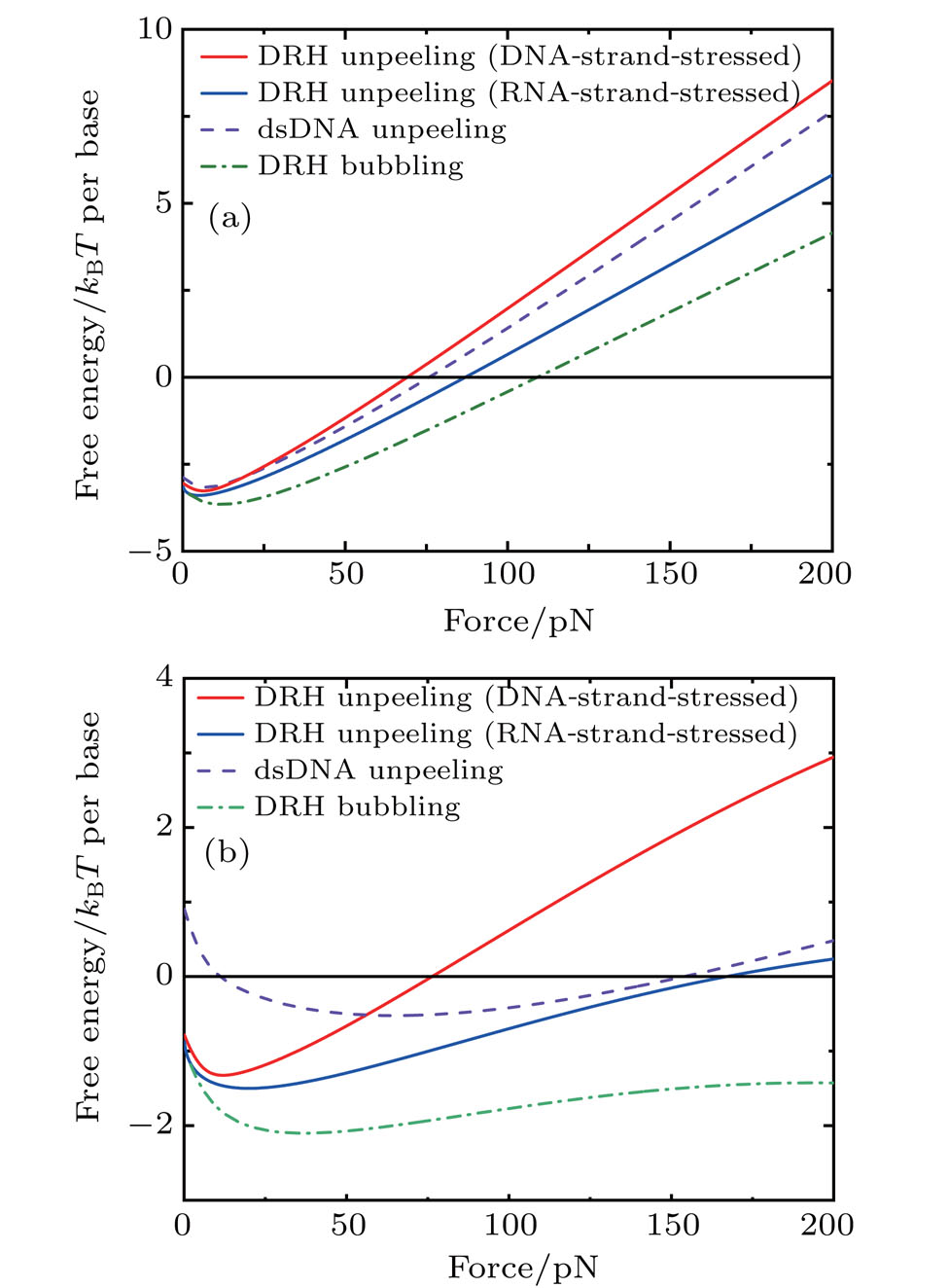
Figure 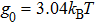



 | Fig. 3. Free energy per base pair of unpeeled and bubbled DRH relative to (a) double-helix form DRH and (b) S-form DRH, with sequence effect averaged out. |
It is also noticed in Fig.
In consideration of the sequence effect, we average the base-pairing free energy of rGG, rGC, rCC, and rCG of Table 



 | Fig. 4. Plots of free energy per base pair of unpeeled and bubbled DRH relative to ((a), (c)) double-helix form DRH and ((b), (d)) S-form DRH for ((a), (b)) extremely GC-rich sequence and ((c), (d)) extremely AT(u)-rich sequence. |
Furthermore, we consider the effects of the purine content on the stability of different forms of DRHs. Previous studies[14] suggested that the general thermal stabilities of the DNA, DRH and RNA duplexes are ordered as 
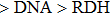


 | Fig. 5. Free energy per base of unpeeled DRH relative to (a) double-helix form and (b) S-form for different pyrimidine content in DNA strand of DRH. |
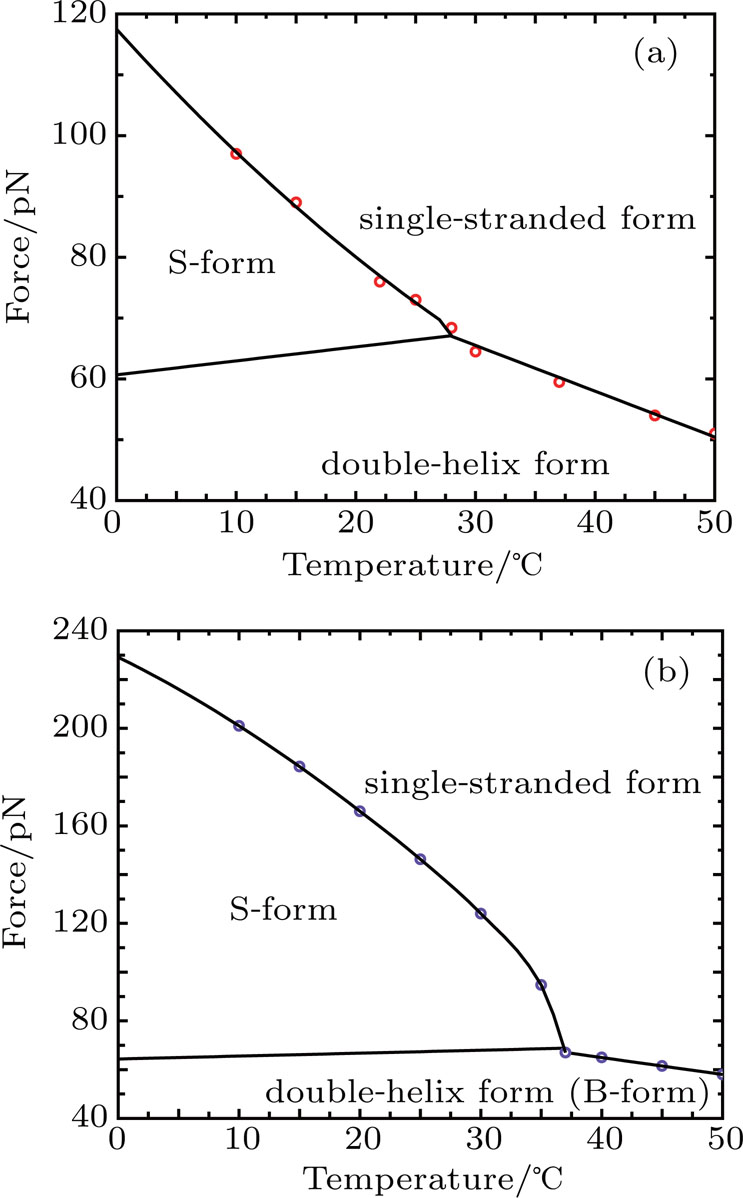
After examining the sequence effect, we further investigate how the temperature affects the stabilities of different forms of DRHs. To show the temperature effect more clearly, we calculate the force-temperature phase diagrams for both DRH and DNA. Here we choose the DNA-strand-stressed DRH as an example because this template is more generally used in the experiments (i.e., in most of experiments, the DNA strand of DRH is often the tethered strand because labeling DNA strand with linkers is much easier than labeling RNA strand). To obtain the force-temperature phase diagrams, we calculate the free energy differences from Eqs. (

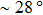
 | Fig. 6. Force–temperature phase diagram of (a) DNA-strand-stressed DRH and (b) double-stranded DNA, both for 50% GC and pyrimidine content under 1-M NaCl condition. Circles denote calculated unpeeling force. |
Finally, to better understand the hysteresis in the force–extension curves measured in DRH stretching/relaxation experiments, we resort to a kinetic model to simulate the stretching and relaxation process of a DNA-strand-stressed DRH molecule with a nick at the end (n0=0). The simulation condition is the same as the above, i.e., at 25 °C under 1-M NaCl buffer condition. Figure 
 | Fig. 7. Simulated stretching and relaxation curves of DRH with a nick at the beginning of the sequence (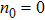
|
Finally, in the simulation, we can also compare the stretching and relaxation processes at different temperatures as shown in Fig.
4. Conclusions
We use a thermodynamic theory to analyze the free energy and competitions among different processes during mechanically stretching a DRH molecule. These results are also compared with those of the DNA.
First, without considering the sequence effect, our calculation shows that the DNA-strand-stressed DRH is easier to unpeel than the dsDNA while RNA-strand-stressed DRH remains, for it is the hardest to be unpeeled in all these three molecules. In addition, the bubbling is always less favored than unpeeling or forming the S-form DRH under all our simulation conditions.
Then in consideration of the sequence effect, our calculation shows that increasing GC content in DRH can significantly increase the stability of the molecule while increasing purine content in the DNA strand of DRH reduces the stability of the molecule. Moreover, for the AT-rich and purine-rich sequence, unpeeling always happens before the transition from the double-helix to the S form, while for the GC-rich and pyrimidine-rich sequence, the opposite scenario happens. In addition, in all cases, the bubbling is always the least favored even for the AT- or purine-rich sequence within our simulated force range.
Our calculation also demonstrates that the temperature plays a significant role in determining the relative stabilities of various forms of DRH and DNA. If the temperature is lower than the triple point in the force-temperature phase diagram of DRH and DNA, as the force increases, both DRH and DNA will change from double-helix form to S form first and then start to be unpeeled later. However, if the temperature is beyond the triple point, both DRH and DNA will not undergo the transition to the S from. Instead, they directly start to be unpeeled from the double-helix form during stretching. It is noticed that the temperature at the triple point for DRH with 50% GC and pyrimidine content is only 28 °C, much lower than that for DNA with the homologous sequence (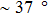
Further, using the kinetic model, we simulate the force–extension response of a DRH molecule during its stretching and relaxation process under different pulling rates and temperatures. Our simulation shows both the pulling rate and the temperature affect the overstretching kinetics significantly. The higher pulling rate generally leads to the larger unpeeling force and overall hysteresis. In contrast, temperature has a more important influence on DRH overstretching kinetics. For example, increasing temperature will greatly lower the unpeeling force. Moreover, because the S-DRH cannot be formed as the temperature exceeds the triple point, the stretching and relaxation process will exhibit different features when the temperature is below or above the triple point.
In summary, our work provides a detailed description of the thermodynamic and kinetic properties of the overstretching transitions of DRH. The calculated results also allow direct comparisons with single molecule stretching experiments. We thus expect it to be a useful guide for the future study of the overstretching thermodynamics and kinetics of DRH under various conditions. The results here may also help gain an insight into other DRH-associated biological processes, such as the R-Loop formation during transcription.[31,32]
Acknowledgment
We want to thank Prof. Jie Yan in National University of Singapore for very helpful discussion.
Reference
| [1] | |
| [2] | |
| [3] | |
| [4] | |
| [5] | |
| [6] | |
| [7] | |
| [8] | |
| [9] | |
| [10] | |
| [11] | |
| [12] | |
| [13] | |
| [14] | |
| [15] | |
| [16] | |
| [17] | |
| [18] | |
| [19] | |
| [20] | |
| [21] | |
| [22] | |
| [23] | |
| [24] | |
| [25] | |
| [26] | |
| [27] | |
| [28] | |
| [29] | |
| [30] | |
| [31] | |
| [32] |