

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Modulation and control of DNA charge inversion
Cite this Article
Wang Yan-Wei, Yang Guang-Can. Modulation and control of DNA charge inversion. Chinese Physics B, 2017, 26(12): 128706 
Permissions
Modulation and control of DNA charge inversion
† Corresponding author. E-mail:
Abstract
DNA is one of most important biological polyelectrolytes, which is negatively charged in physiological condition. Most of Its charge is neutralized by attracting cations in solution. In some conditions, the effective charge of DNA switches its sign from negative to positive, implying charge inversion of DNA. The underlying microscopic mechanism of the counterintuitive phenomenon is still not fully understood although specific chemical affinity and electrostatic ion correlation are considered as two possible driving forces. In this review, we present some recent experimental progress in the modulation and control of DNA charge by single molecular techniques. It has been shown that DNA charge inversion can be modulated bidirectionly by decreasing or increasing the dielectric constant of solution to make the electrophoretic mobility of DNA increase from a negative value to a positive value. In this meanwhile, charge inversion and condensation of DNA in solution of trivalent and quadrivalent counterions are significantly influenced by pH value of the solution. When mixing quadrivalent counterion with mono-, di- and tri-valent counterions in solution, suppression and promotion of DNA charge inversion can be observed. In addition, hydrophobic effect can play an important role in DNA charge inversiton and compaction. We show that the organic monovalent ions of tetraphenyl chloride arsenic (Ph4As+) can induce DNA compaction and even invert its electrophoretic mobility. Thus, hydrophobic effect can be the main driving force of DNA charge inversion and compaction by the organic monovalent ion.
1. Introduction
DNA, as an important biological polyelectrolyte, is highly charged when dissociated in solution and surrounded by counterions carrying positive charges. Interactions between DNA and mobile cations in solution play an important role in DNA physical properties and biological function.[1,2] The complicated interactions lead to two counterintuitive phenomena:like-charge attraction between macroions and charge inversion (CI), in which a macroion attracts more opposite charges in excess of its own nominal charge so that its effective charge changes its sign. For DNA system, the like-charge attraction results in its compaction and condensation, which enable it to be highly packaged in various organisms from viruses to eukaryotic cells in order to store, transport and preserve the genetic material. Thus, the underlying mechanism of charge inversion and condensation of DNA induced by counterions is not only important for understanding basic biological process, but also helpful in developing new techniques for DNA extraction, gene therapy and gene transfection.[3] CI and condensation of DNA depend on many factors, including valance state, concentration and type of counterions. Over the years, a number of experimental and theoretical studies have been carried out with the goal of providing a general physical picture of the charge inversion and condensation phenomenon of anionic DNA and RNA molecules,[4–13] such as charge-regulated DNA compaction, ion-mediated DNA–DNA interactions, resisting condensation of RNA, and DNA end-to-end attraction.
In recent years, some new single molecular approaches have been used to explore the issue to get more insights. For example, Lemay group[14] observed the reversal of the polarity of charged surfaces in water upon the addition of trivalent and quadrivalent ions by atomic force microscopy. They found that the bulk concentration of multivalent ions at which CI occurs depends almost only on the valence of counterions. Then they turned to DNA system and demonstrated the CI of DNA by multivalent ions through measuring electrophoretic mobility (EM) of condensed DNA by dynamic light scattering (DLS). Magnetic tweezers (MT) is a versatile tool for single molecular force spectroscopy, not only measuring force but also being able to measure torque of tethering molecules.[15] Measuring the multivalent-ion induced condensation of a single DNA molecule by using MT, they further showed that Clis related to the condensation by modulating the barrier for condensate nucleation.[3] For DNA systems, only quadrivalent counterions of high concentration can lead to the CI, which is deviated from the theoretical prediction of strong correlation theory significantly. Recently, optical tweezers (OT) were employed to study the electrophoretic and the electroosmotic motion of a single colloid immersed in electrolyte solutions of different valences, and it is found that at low colloidal charge densities, ion correlation effects alone do not suffice to produce mobility reversal.[16] On the other hand, Lösche group demonstrated the CI of a lipid monolayer at ultralow electrolyte concentrations, much lower than the predicted of ion-ions correlation theories. They suggested that transverse electrostatic correlations between mobile ions and surface charges might play an important role in the process.[17] Using streaming currents, Dekker group observed that CI occurs in rectangular silica nanochannels at high concentrations of divalent ions.[18] Regardless the accumulating observations and theoretical analysis, we need more experimental study to clarify the dominant driving mechanism for CI. Fluorescent microscopy and single molecule electrophoresis.[19]
CI of DNA has been theoretically predicted by strong correlation theory, and confirmed by numerical simulation in which they clarified the difference between the inversions of DNA’s electrical charge and its electrophoretic mobility.[20] CI of DNA induced by counterions is also characterized through extensive molecular dynamics simulations.[21] The simulation clarifies the difference between the inversion of the electric charge and the electrophoretic motion of DNA resulting from a complex interplay of electrostatics and hydrodynamics. The effect of dielectric constant and finite ion size also modify ion-mediated forces between DNA molecules and are quantitatively investigated by the Monte Carlo method.[22] Although both strong correlation theory and numerical simulation predict the CI of DNA upon the high concentration of trivalent or quadrivalent counterions, it has not been observed for DNA in trivalent solution regardless its high concentration.
In the present review, we present some experimental progress about DNA charge inversion and compaction. DNA CI can be modulated by adjusting the dielectric properties of solutions.[23] We can make CI happen in trivalent counterion solution by adding ethanol to decrease the dielectric constant of medium, which is realized in trivalent counterion solutions. Meanwhile, we can also show the opposite process to neutralize DNA CI by adding zwitterions to the solution of counterions having 4 or more valences, so to raise its dielectric constant. It is remarkable that adjusting the dielectric constant of the medium can be achieved in two ways through adding ethanol or zwitterionic species (ZS) to the solution of DNA. Dielectric constants of the solution also modulate the condensing and unraveling forces significantly.
In a mixed ion solution containing a mild amount of tetravalent cations,[24] charge neutralization of DNA is suppressed by mono- and divalent cation but promoted by trivalent cations. On the other hand, pH value in solution influences on the condensation and charge neutralization of DNA.[25]
In the case of monovalent ions, electrostatic correlations are weak and no CI is expected. However, if hydrophobic effect is dominant, it can be the main driving force to DNA CI and compaction. For example, when introducing the organic monovalent ion Ph4As+ into a DNA system and it is found that it not only provokes the CI of DNA, but also leads to DNA compaction.[26] This phenomenon seems to be similar to the poor solvent effect of neutral condensing agents such as ethanol in DNA solution.[27]
2. Experimental methods
The experimental techniques used to study DNA CI and compaction include DLS, single molecular electrophoresis (SME), atomic force microscopy (AFM),[28] and MT. Among then, DLS is a feasible tool for electrokinetic properties of DNA and its complexes.
Light scattering is a consequence of the interaction of light with small particles or molecules in medium. When particles become larger than the tenth of wavelength of light, the scattering changes from being isotropic to a distortion in the forward scattering direction. From the signal of DLS, we can extract speed information of particles undergoing Brownian motion, influenced by particle size, medium viscosity, and temperature. According to Stokes–Einstein theory, we can calculate the particle size from these measured quantities. When an oscillating electric field is applied to the sample, we can obtain additional electrokinetic properties such as zeta potential and electrophoretic mobility of particles.
In SME experiment, the DNA mobility was measured by recording DNA migration video in fluorescent microscopy. AFM is used for the observation of the morphologies of DNA complexes and MT for measurement of condensing and unraveling forces of single DNA molecule pulling.
EM measurements were carried out using two methods. One was carried out by using DLS device of Malvern Zetasizer nano ZS90 equipped its patented M3-PALS technique, in which an He–Ne gas laser (λ = 633 nm) is used and the light scattering is recorded by the avalanche photodiode mounted on the goniometer arm at to the direction of the incident radiation. In our experiment, 1-ml DNA solution sample was placed in the folded capillary cell for size and electrophoretic mobility measurement.
Another EM measurement was by SME. It is based on an inverted fluorescence microscope (Nikon, TE-2000E) equipped with an oil immersion objective (Nikon, 100X, N.A. = 1.49) and an ND filter slider (Nikon, 330–385/460–490/510–550 nm). A 100-W high pressure mercury lamp served as illumination source. The intensified charge-coupled device (CCD) camera (512×512 pixels of Cascade II512) was used for video recording, which was used for analyzing DNA EM. In the electrophoresis, the DNA was stained with YOYO-1 fluorescent dye at a dye-base-base ratio of 1:10 with TRIS buffer before using.
A custom-made transversal MT is used for condensing and unraveling forces of tethered DNA molecules.[26] In the experiment, a DNA-bead construct was rinsed into the flow cell to form a side wall-DNA-paramagnetic beadstructure. A magnet is applied to pull the beads. The distance between the bead and the surface of the sidewall can be measured as the extension of DNA. The applied force was calculated according to Brownian motions of the beads in the direction perpendicular to the DNA extension.
The AFM imaging was performed with a multi-mode AFM with nanoscope controller (SPM-9600, Shimadzu, Kyoto,Japan) in the tapping-mode. All AFM images were captured at conventional ambient tapping mode.
3. Modulating charge inversion of DNA by dielectric properties
In a high-concentration multivalent electrolyte, inversion of the DNA’s electric charge[29] can be directly observed through the motion reversal of DNA in solution when an external electric field is applied. Electrophoretic mobility (EM) of DNA, μ, relating with the net charge of DNA and the counterions condensed at its surface can be measured by DLS in solution.
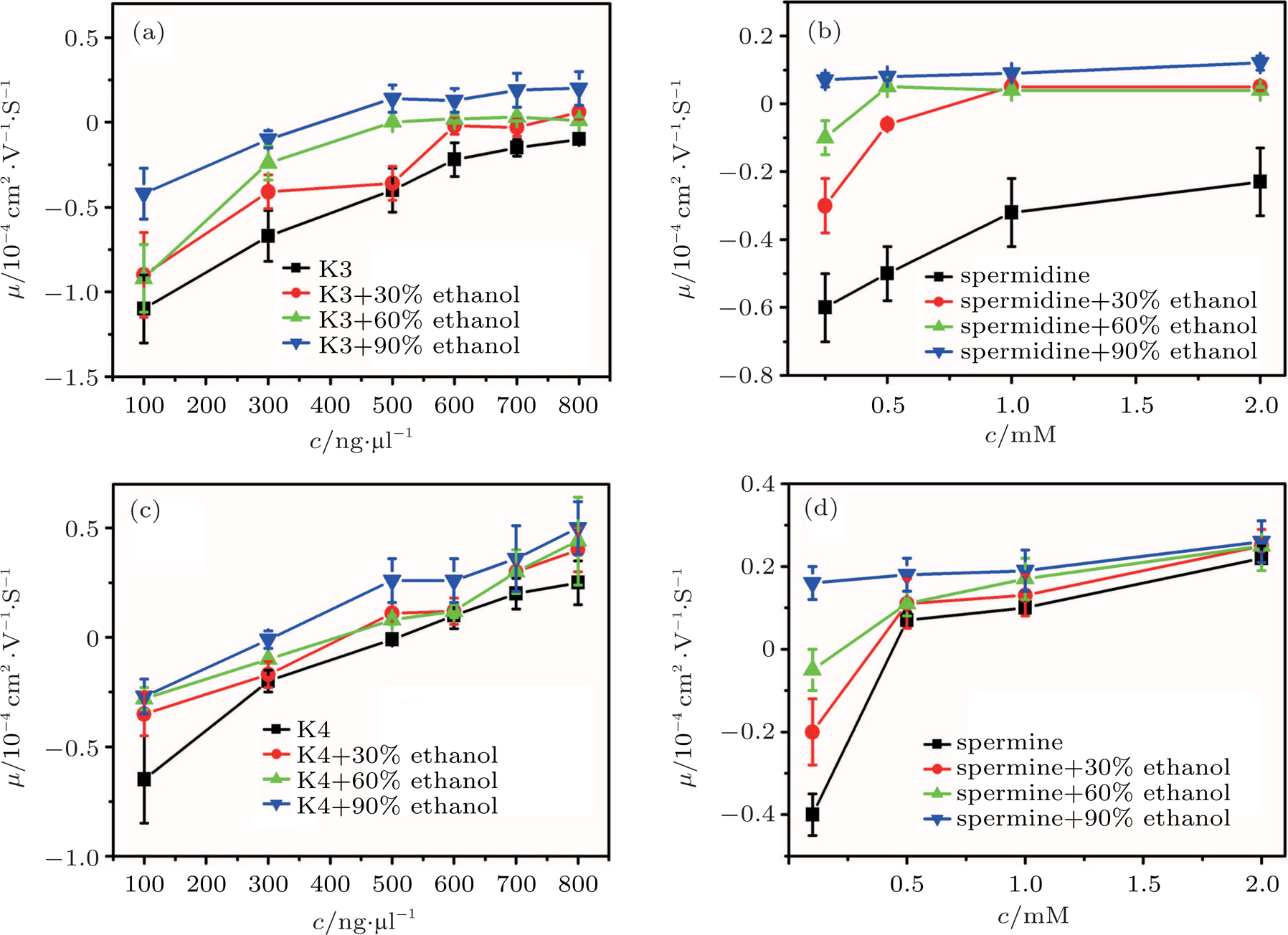
The electrokinetic properties of DNA in trivalent and quadrivalent ion solution are shown in Fig.
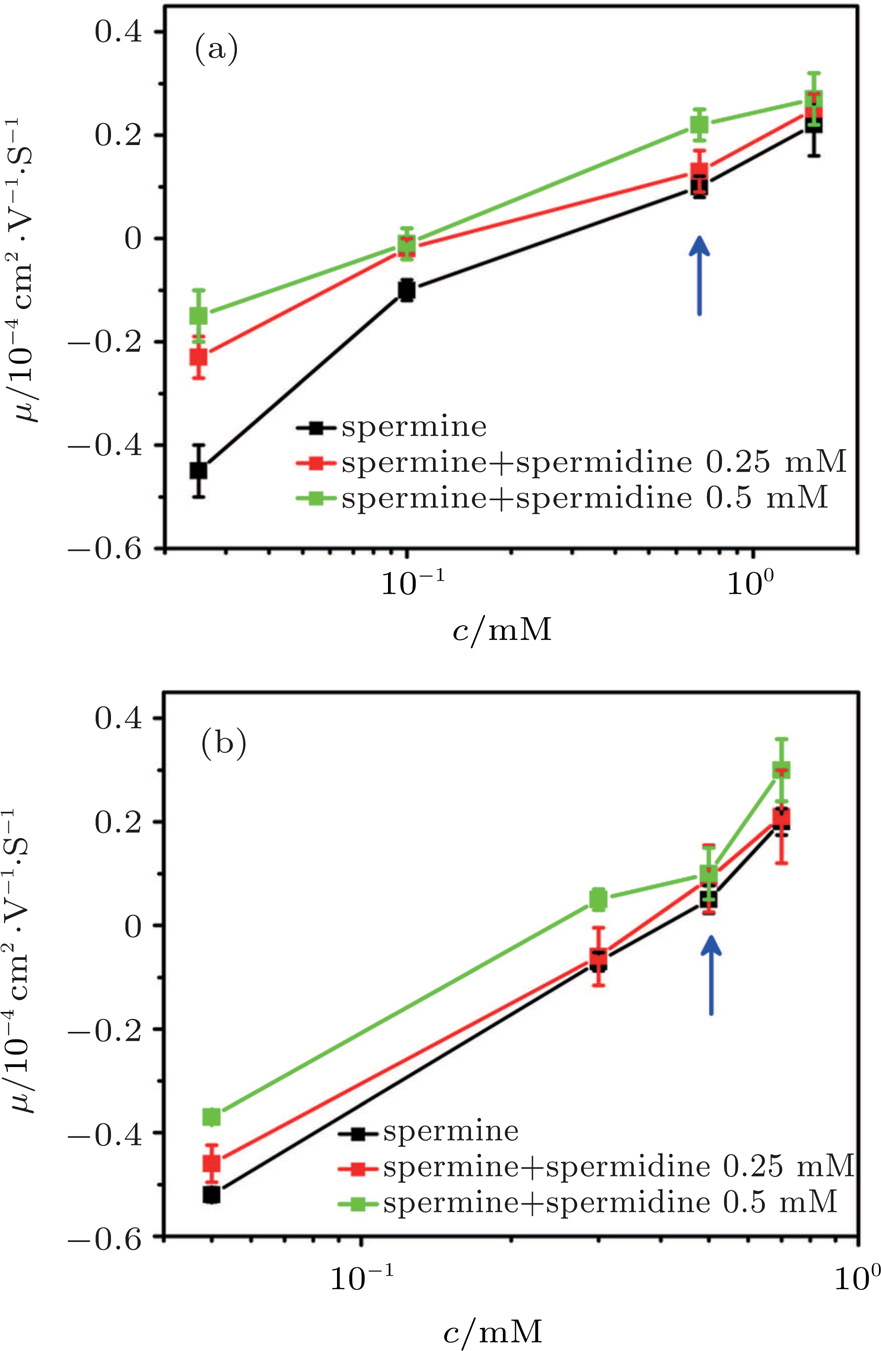
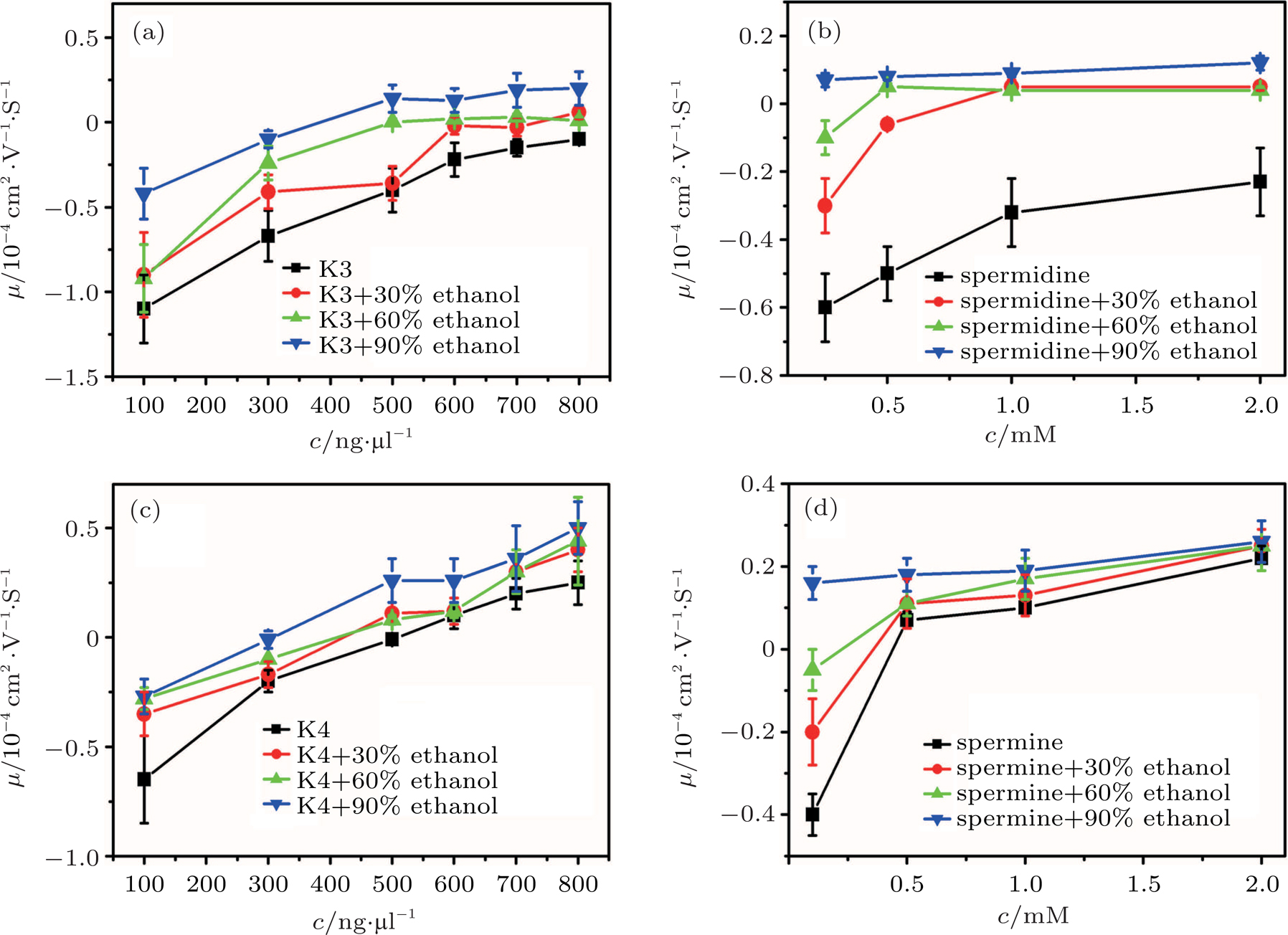 | Fig. 1. (color online) Electrophoresis mobility μ, of condensed DNA as a function of K3, K4, spermine, and spermidine concentration in 1-mM TRIS and various mixtures of ethanol. |
As for quadrivalent K4 and spermine, EMs of DNA become positive from a negative value at critical concentrations of 0.5 mM and 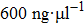
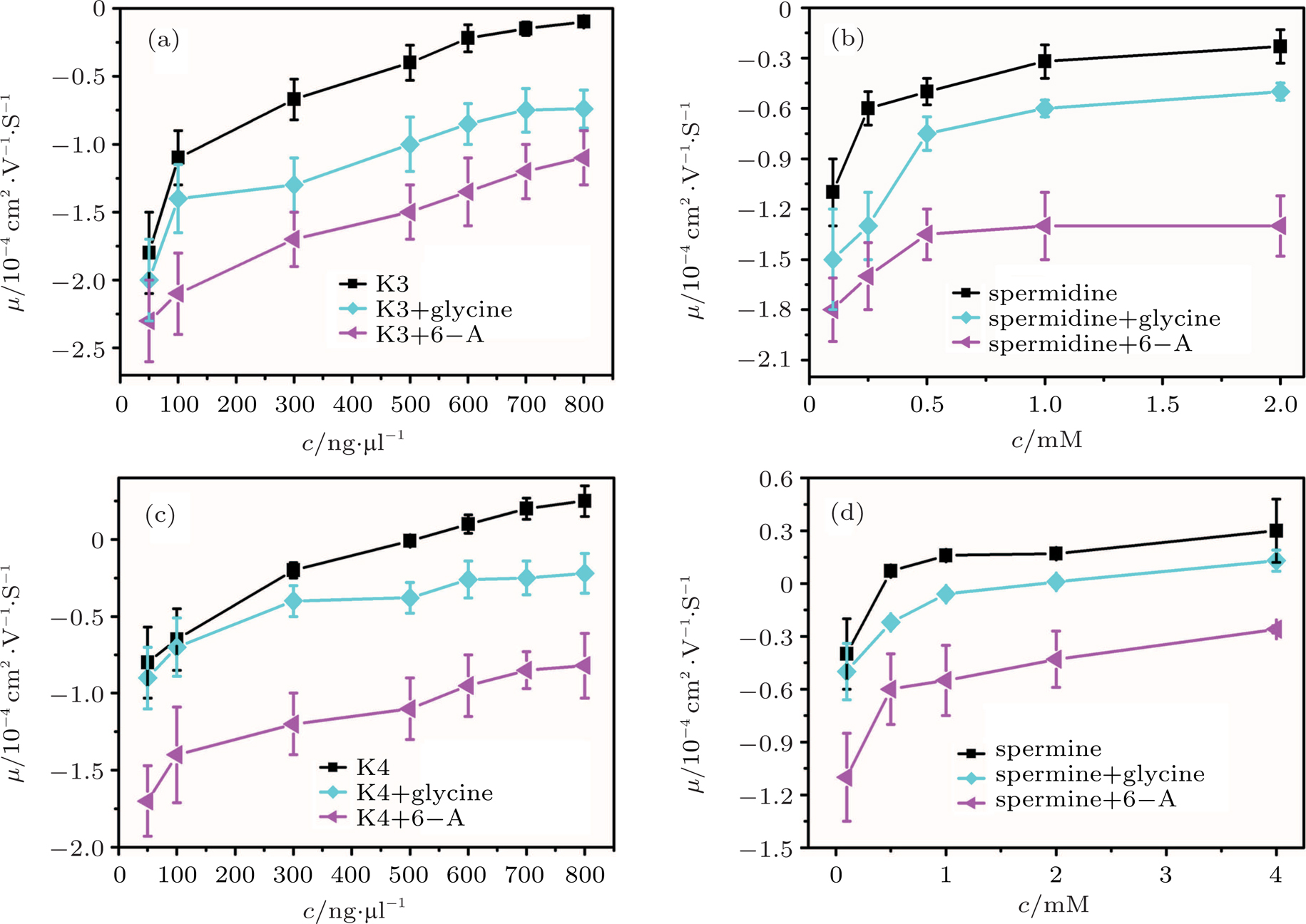
In other words, lowering dielectric constant of medium make CI of DNA occur more easily in the same ion environment. The observation implies that the driving force of DNA CI is electrostatic interaction because of only adjusting dielectric constant. To rule out the specific interaction between ethanol and DNA, we can execute a similar procedure but increasing the dielectric constant of solution. To do so, we add different glycine, 6-A to the aqueous solution so that the dielectric constant of medium has an increment depending on the concentration of these amino-carboxylic acids. The measured EM by DLS is shown in Fig.
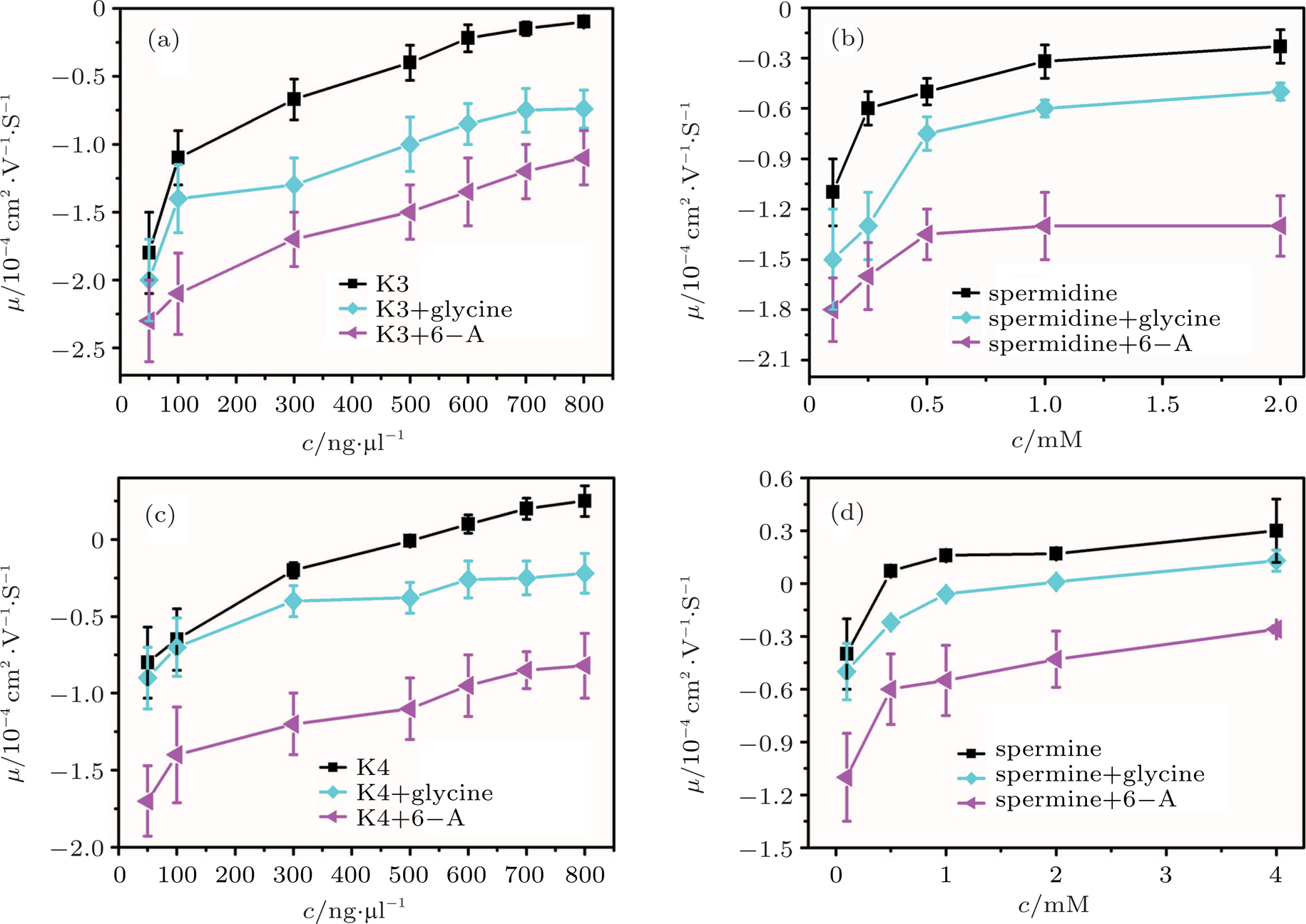 | Fig. 2. (color online) Electrophoresis mobility, μ, of condensed DNA as a function of K3, K4, spermine, and spermidine concentration in 1-mM TRIS with various zwitterions. |
CI and like-charge attraction are two distinct counterintuitive phenomena of electrostatics in solution, and are closely relevant. The latter play a significant role in various types of macromolecular organization in charged systems, from clustering in colloidal suspensions to precipitation offlexible polyelectrolytes.[30] For DNA system, like-charge attraction leads to DNA compaction or condensation, in which condensing force is closely related to its charge neutralization. Since Lemay’s group investigated the folding and unfolding processes of single double strand DNA chains undervarious ionic conditions, we focus on the role of dielectric property of solution.
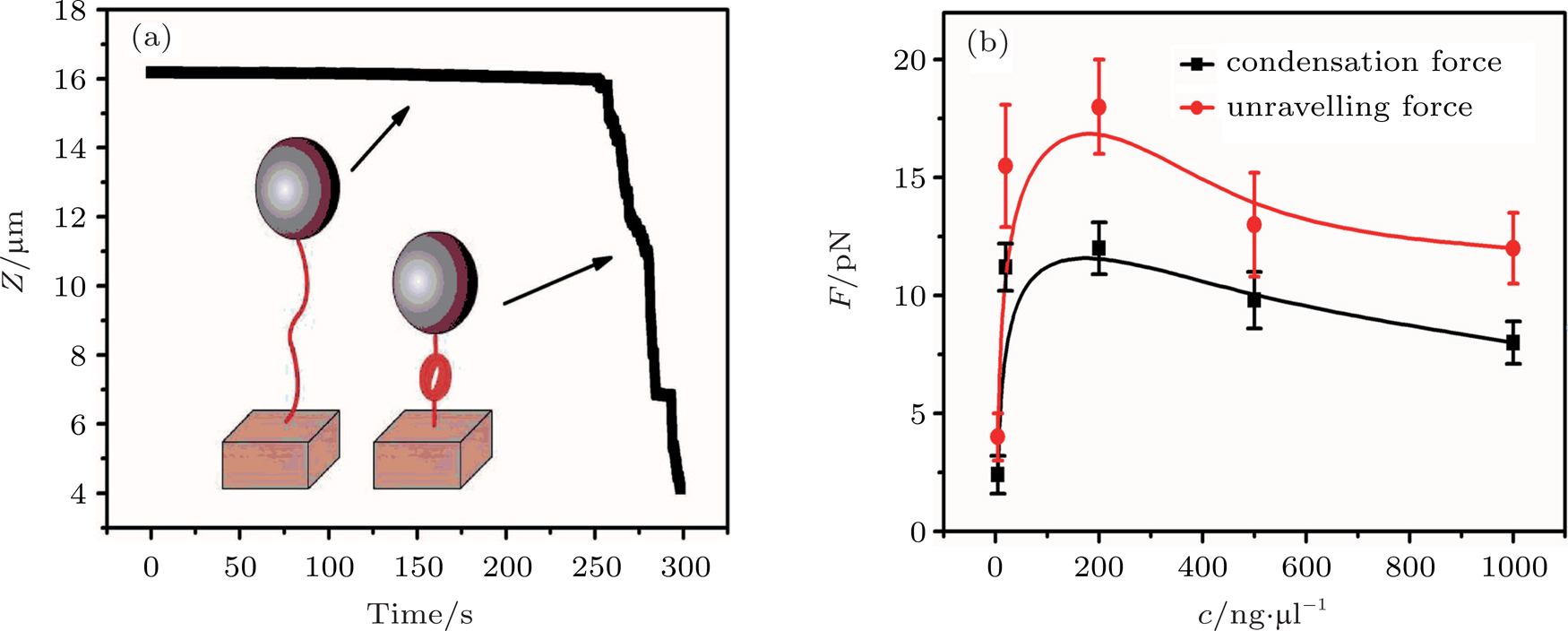
By MT, we can tether DNA compaction and measure the applied force simultaneously. In the setup, condensing force (

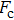
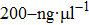
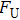
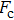
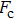
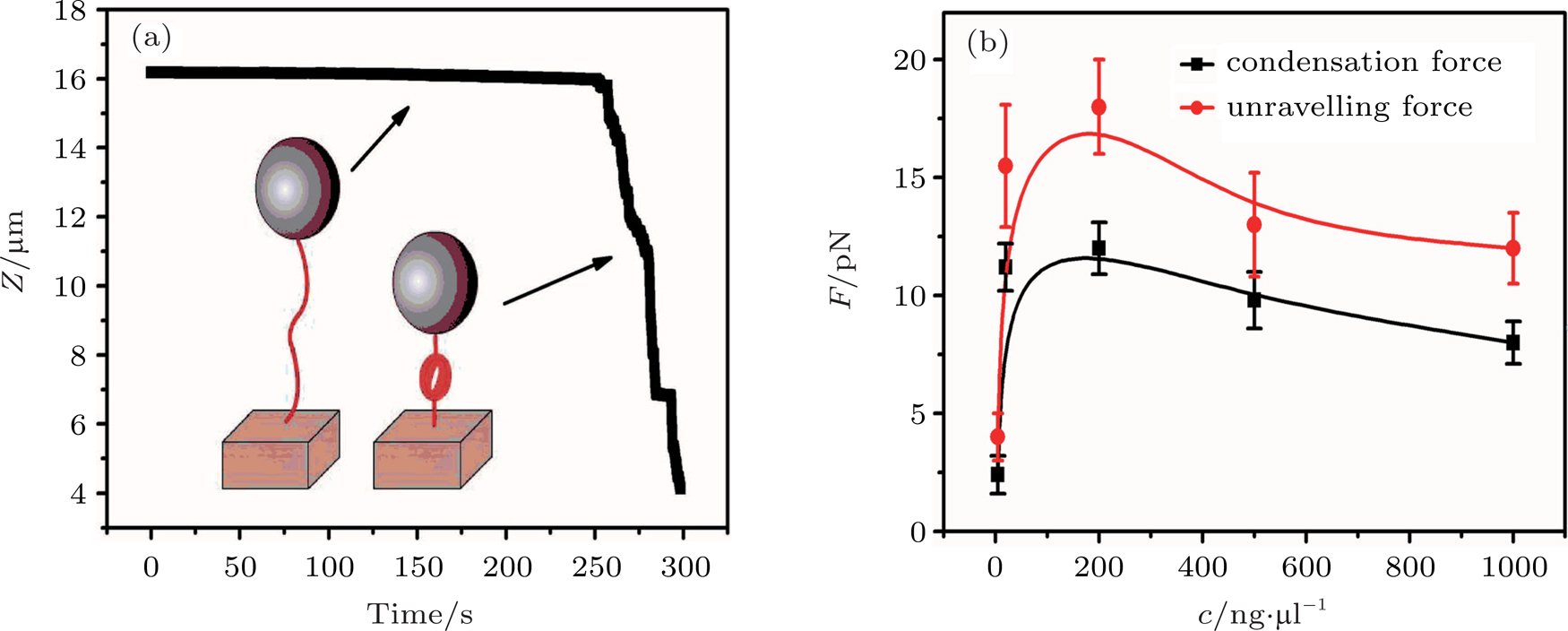 | Fig. 3. (color online) The curve of condensation, condensing, and unraveling forces. (a) DNA extension–time curve measured by MT in DNA condensation process with K8. (b) 

|
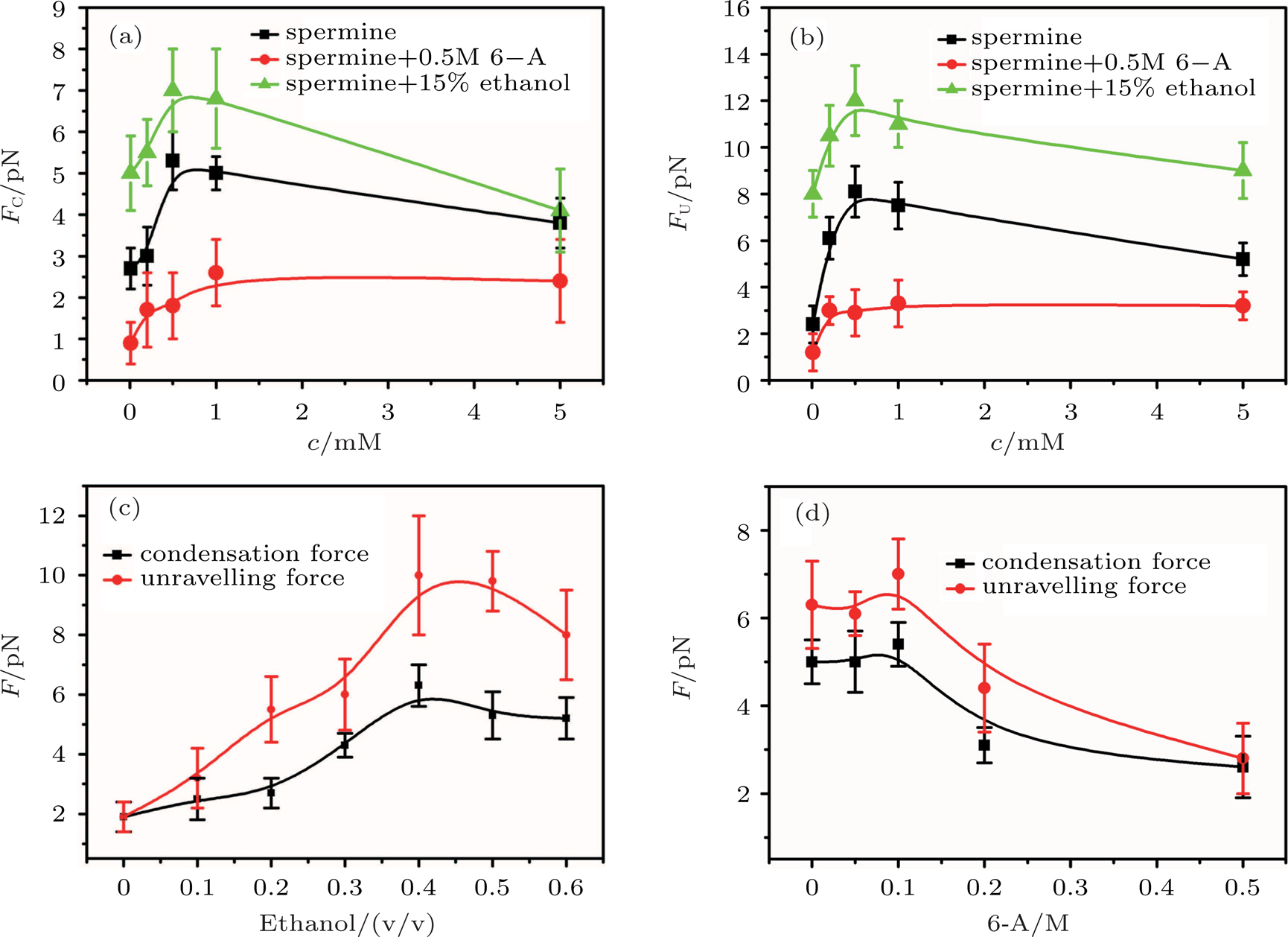
In general, the DNA pulling shows that the maximal condensing or unraveling force corresponds to the completely neutralization of DNA charge, under or over compensation of charge results in descending of the force due to the enhancing Columbic repulsion. As shown in Fig.
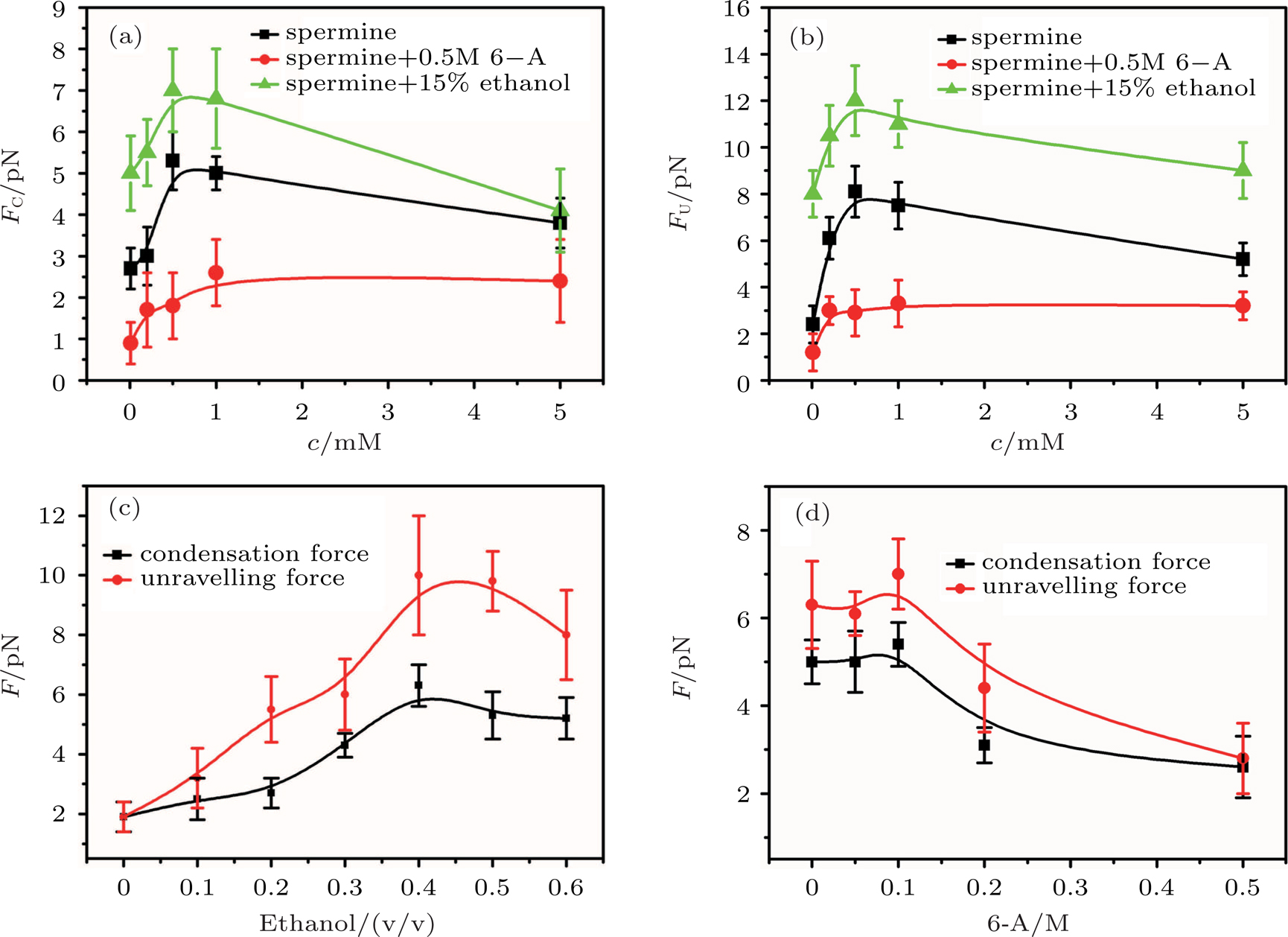 | Fig. 4. (color online) 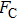
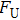
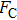
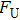
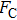
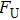
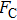

|
In the framework of Poisson–Boltzmann equation, most of the charge of macromolecules can be neutralized by counterions in solution.[31] However, CI cannot be explained by conventional mean-field theories of charge screening. Among the possible mechanisms of specific chemical adsorption and strong correlation effect, based on the experimental data here, we can draw a conclusion the CI in DNA system is driven predominantly by correlated electrostatic interactions. Although the dielectric constant seems to be the main parameter that controls the CI of DNA, one should discuss the eventual effects of the specific interaction between solvents and DNA. The local dielectric constant around the chain is thus expected to be different from that in the bulk solution. In the case of alcohols, Rau group have also shown that such exclusion mechanisms can affect the DNA precipitation by multivalent counterions such as spermidine.[32] Similarly to glycine betaine, the distribution of the zwitterions used in this study around DNA is probably not homogeneous, and exclusion from the DNA chain may be expected. However, by the use of various solvent mixtures, it have been shown that the dielectric constant of the solvent is the key factor that determines the conformational behavior of single DNA molecules in solution.[33] It is reasonable to assume the conclusion is also appropriate for the electrostatic interactions of DNA in solution. However, we only consider the contribution of dielectric properties for simplicity, even if other effects, such as preferential hydration, influence the interactions between solvents and DNA. As pointed by Todd,[34] a change in dielectric constant or solute exclusion are intertwined.
4. Charge inversion and condensation of DNA modulated by pH value
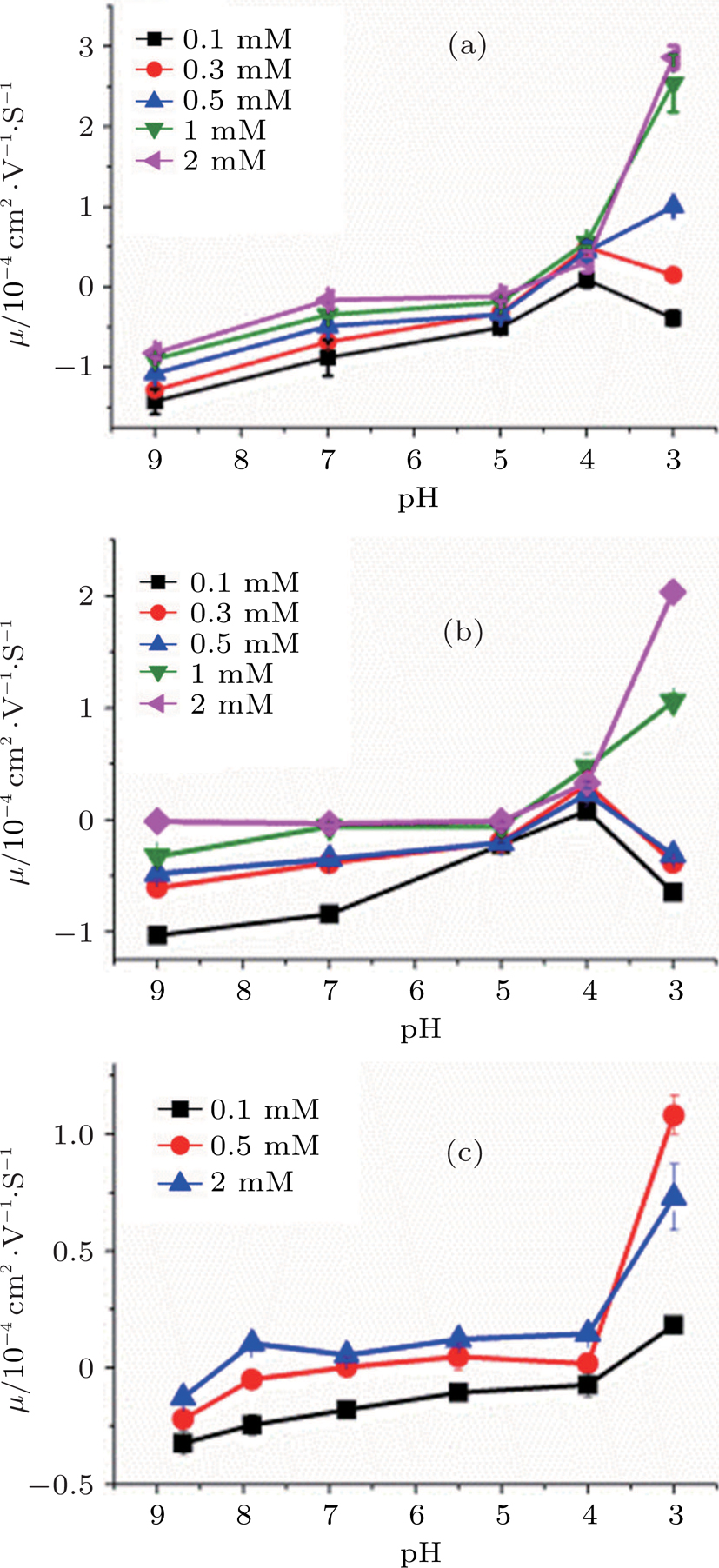
Hydrogen ion (H+) is a special kind of monovalent ion characterized by pH value. It plays an important role in interactions between polymers or colloids and cations, and also has an impact to CI and condensation of DNA. Figure
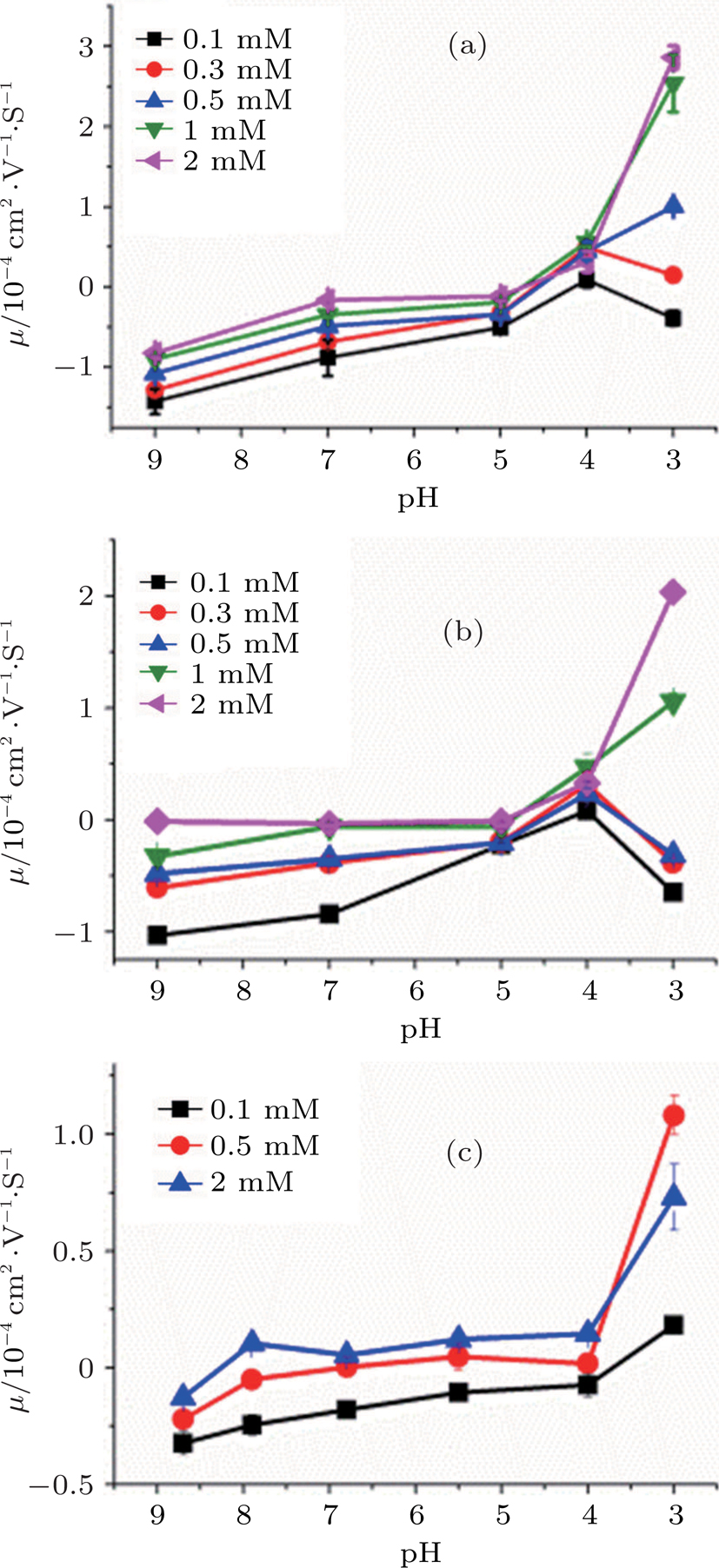 | Fig. 5. (color online) DNA molecule electrophoresis mobility, μ, as a fuction of the pH of solution with TIRS 1 mM in different concentrations of cations. (a) DLS for spermidine, (b) DLS for cobex, the concentration of the solution is 0.1 mM (black), 0.3 mM (red), 1 mM (blue), 0.5 mM (green), 2 mM (pink); (c) DLS for spermine, 0.1 mM (black), 0.5 mM (red), 2 mM (blue). |
Effective valence mechanism of couterions in solution can be applied to the observed variation of DNA charge and electrophoretic mobility. The effective valence of couterions is affected with the concentration of H+. Since polyamines are positively charged with its weak association with H+, they become less charged when the concentration of H+ is lowered, or the pH of solution increases. In other words, the effective valence of the couterions is lowered. For example, the valences of spermidine and spermine in neutral buffer are +3 and +4 respectively. If pH value increases, their effective valences become less than +3 or +4. Accordingly, their ability to neutralize DNA charge is in turn lowered, leading to the electrophoretic mobility of DNA shifts toward the negative region. In contrast, when the concentration of hydrogen ions in solution increases, such as by adding chloride, the effective valence of polyamines increases to recover their ability to neutralize negatively charged DNA.
The effective valence mechanism is most sensitive for polyamines as shown in Fig.
It is noticeable when the concentration of counterions is under the critical value and at very low pH (for example, less than 4), the electrophoresis mobility of DNA decreases rather than increases with the increasing concentration of H+. The phenomenon might be related with DNA denaturation at very low pH, in which DNA may depurinate and melt the double helix, even more, the phosophodiester backbone might hydrolize.[35] However, the descent of mobility does not occur in the cases when the concentration of couterions is above the critical value, in which DNA has been condensed tightly so that the denaturation of DNA is negligible.
Charge neutralization of DNA is also related with its condensation. Figure
 | Fig. 6. (color online) AFM images of DNA with spermine and spermidine in 1-mM TRIS. The inset bar are 
|
5. Suppression and promotion of DNA charge inversion by mixing counterions
When more than one counterionexist in solution, the electro- static interaction between DNA and counterions becomes more complicated. It involves the competition between different species of couterions.
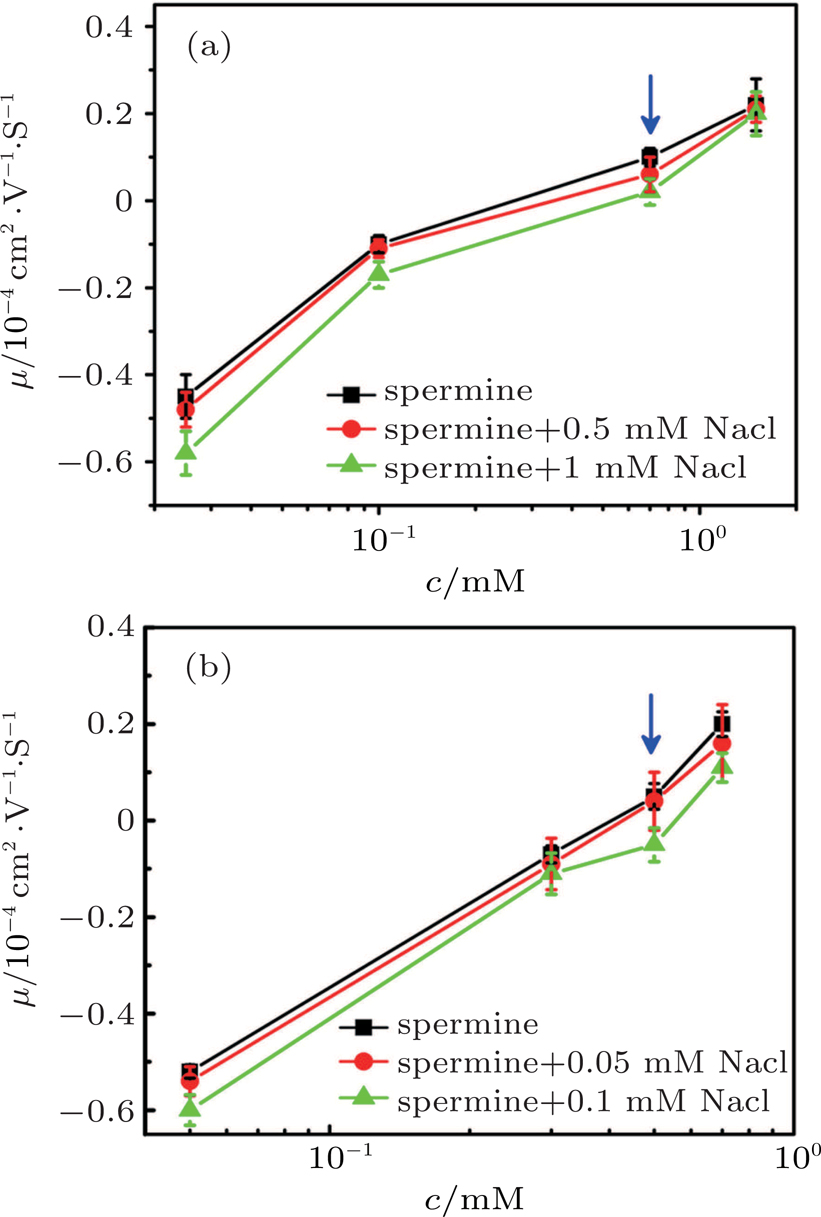
A typical case of mixing counterions is shown in Fig. 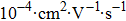
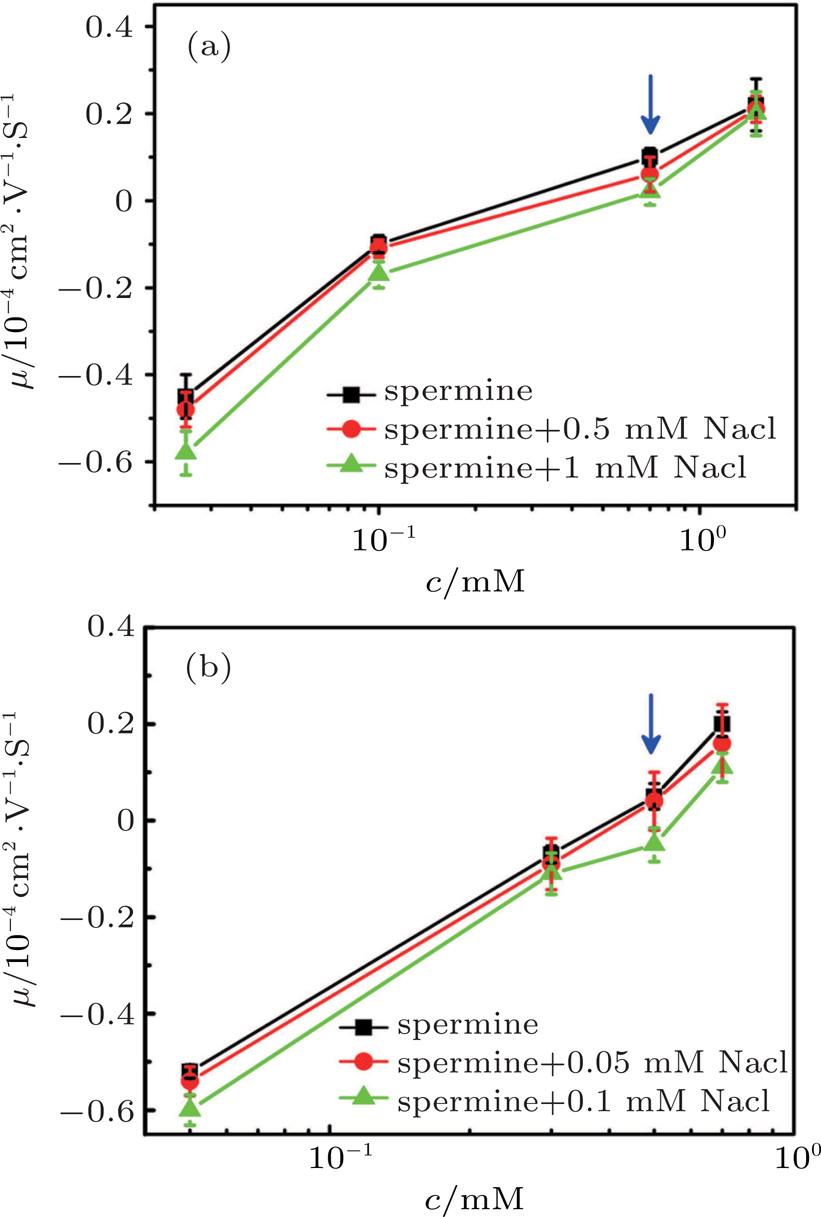 | Fig. 7. (color online) DNA molecule electrophoresis’ mobility as a function of the spermine concentration with monovalent ions. (a) DLS measurements for 0, 0.5 mM, 1 mM sodium chloride (black, red, green). (b) SME measurements for 0, 0.5 mM, 1 mM sodium chloride (black, red, green). The arrows indicate increasing concentration of the sodium chloride. |
To justify the conclusion further, we measured the DNA electrophoresis mobility with 1-mM TRIS buffer containing mixed counterion (NaCl) by SME. The data are shown in Fig.
In Fig.
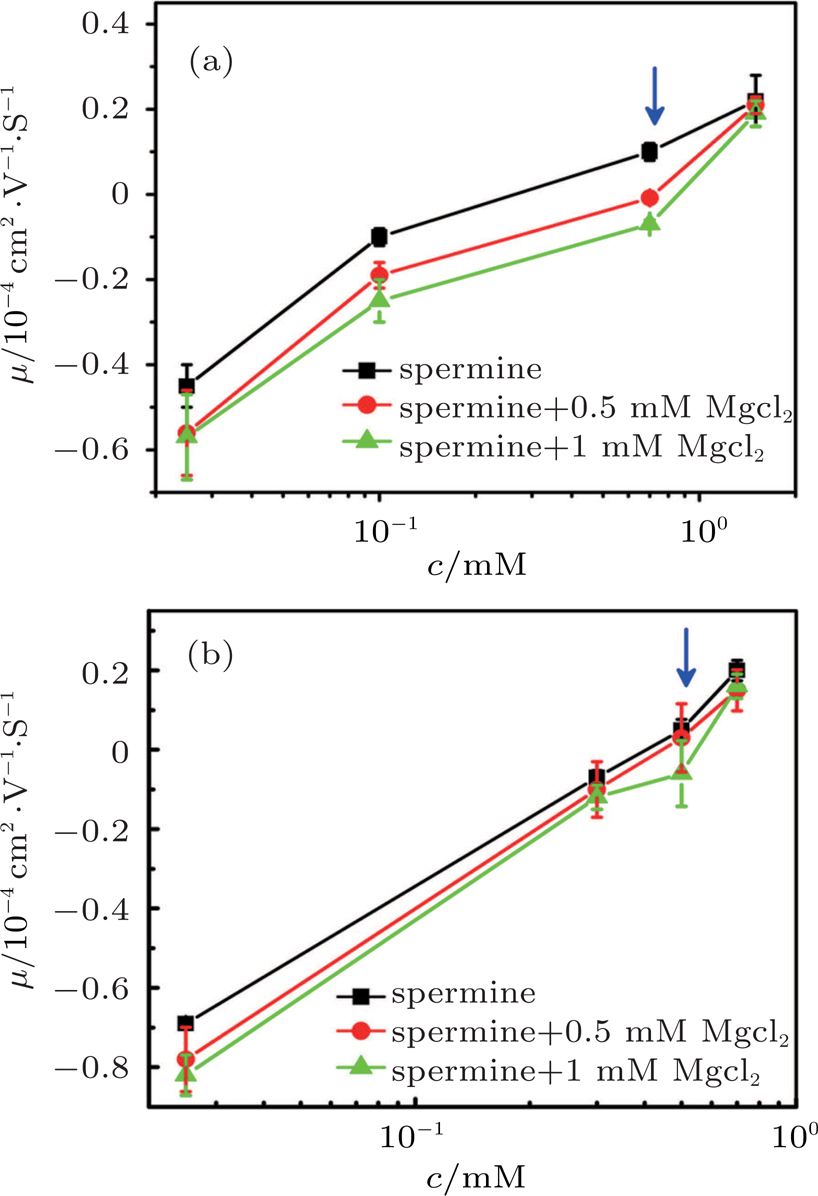
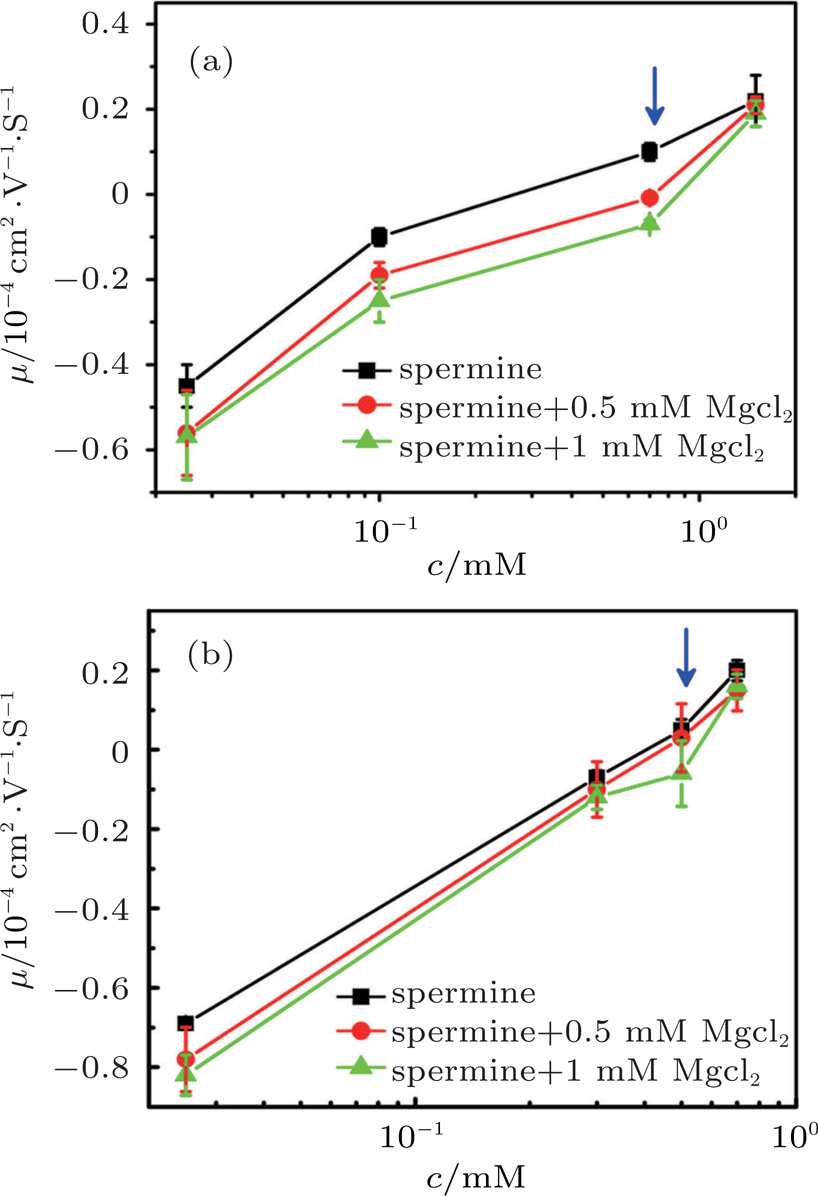 | Fig. 8. (color online) DNA molecule electrophoresis’ mobility μ as a function of the spermine concentration with divalent ions. (a) DLS measurements for 0, 0.5 mM, 1 mM magnesium chloride (black, red, green). (b) SME measurements for 0, 0.5 mM, 1 mM magnesium chloride (black, red, green). The arrows indicate increasing concentration of the magnesium chloride. |
The suppressing effect for the DNA CI depends on the concentration of magnesium chloride. From Figs.
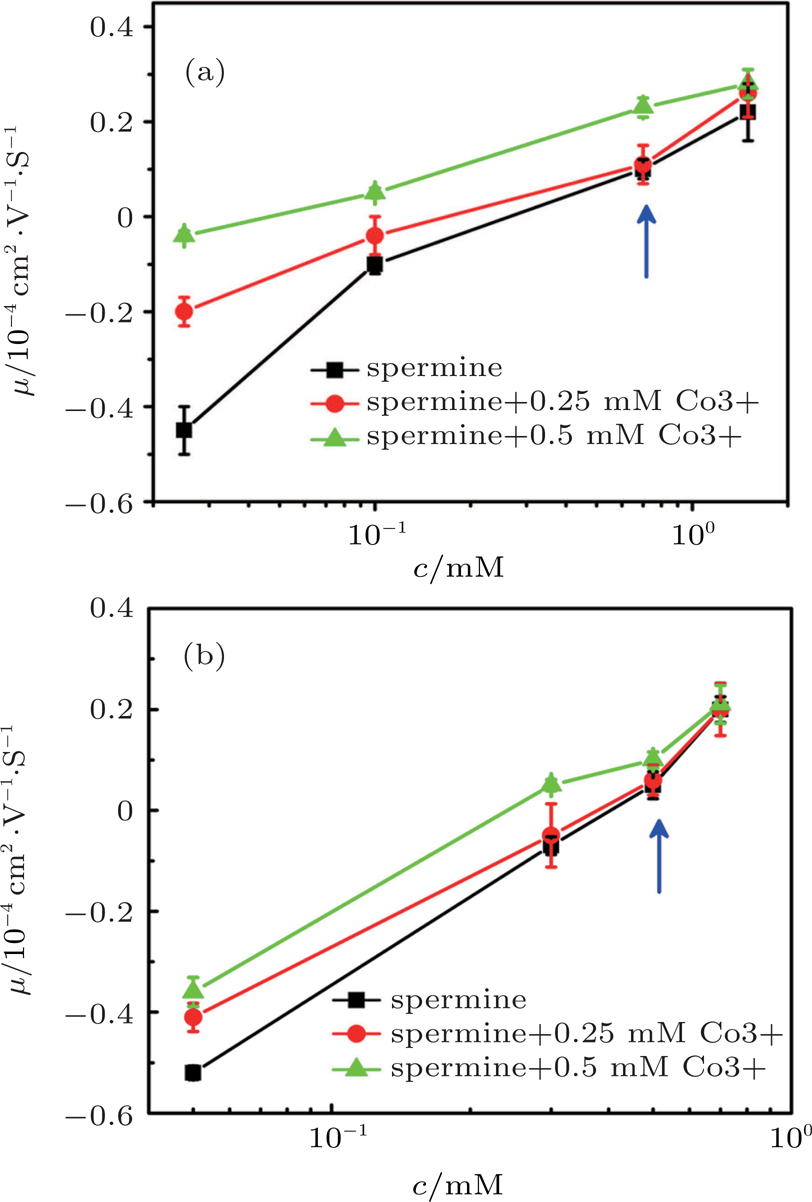
When we add trivalent ions into the solution, some interesting phenomenon happens. Figure
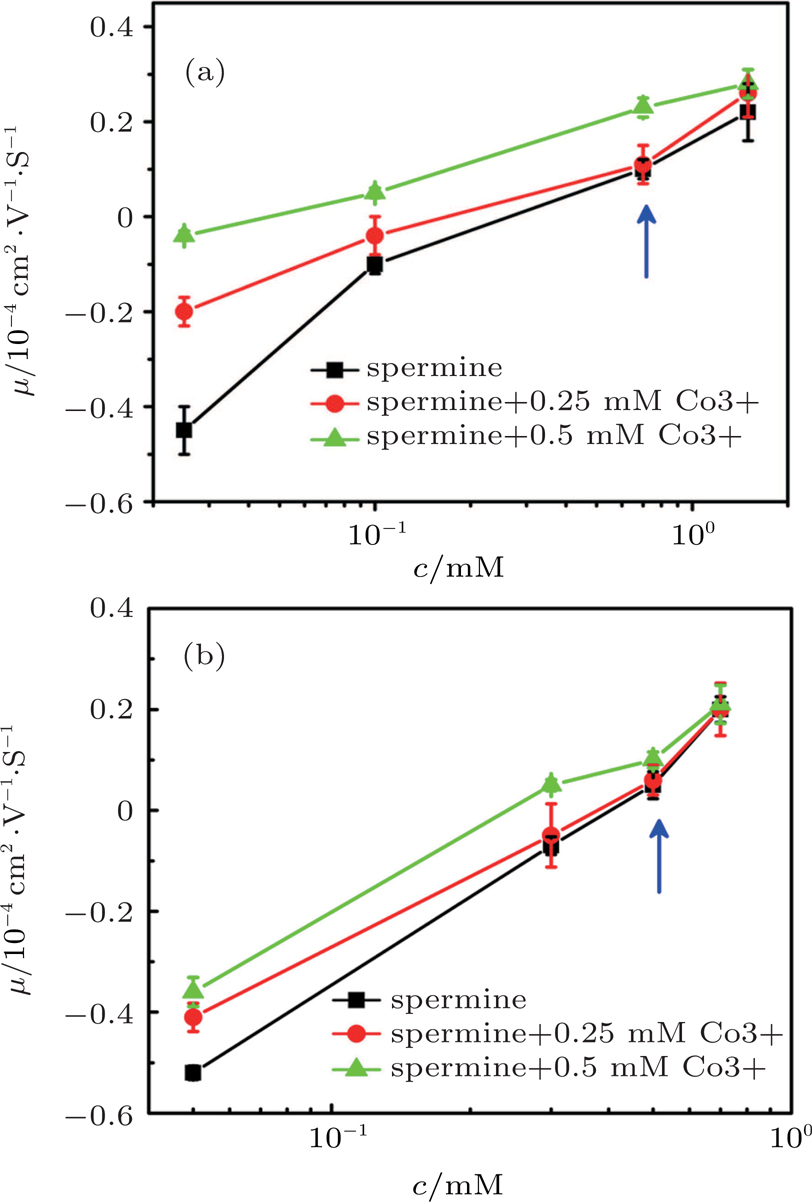 | Fig. 9. (color online) DNA molecule electrophoresis’ mobility μ as a function of the spermine concentration with trivalent cobalt ions. (a) DLS measurements for 0, 0.25 mM, 0.5 mM of hexammine cobalt (black, red, green); (b) SME measurements for 0, 0.25 mM, 0.5 mM of hexammine cobalt (black, red, green). The arrows indicate increasing concentration of the hexammine cobalt. |
To confirm the discovery, we added a different trivalent ion, spermidine, into the spermine and DNA solution to measure DNA electrophoresis mobility. The result from both DLS and SME is shown in Fig.
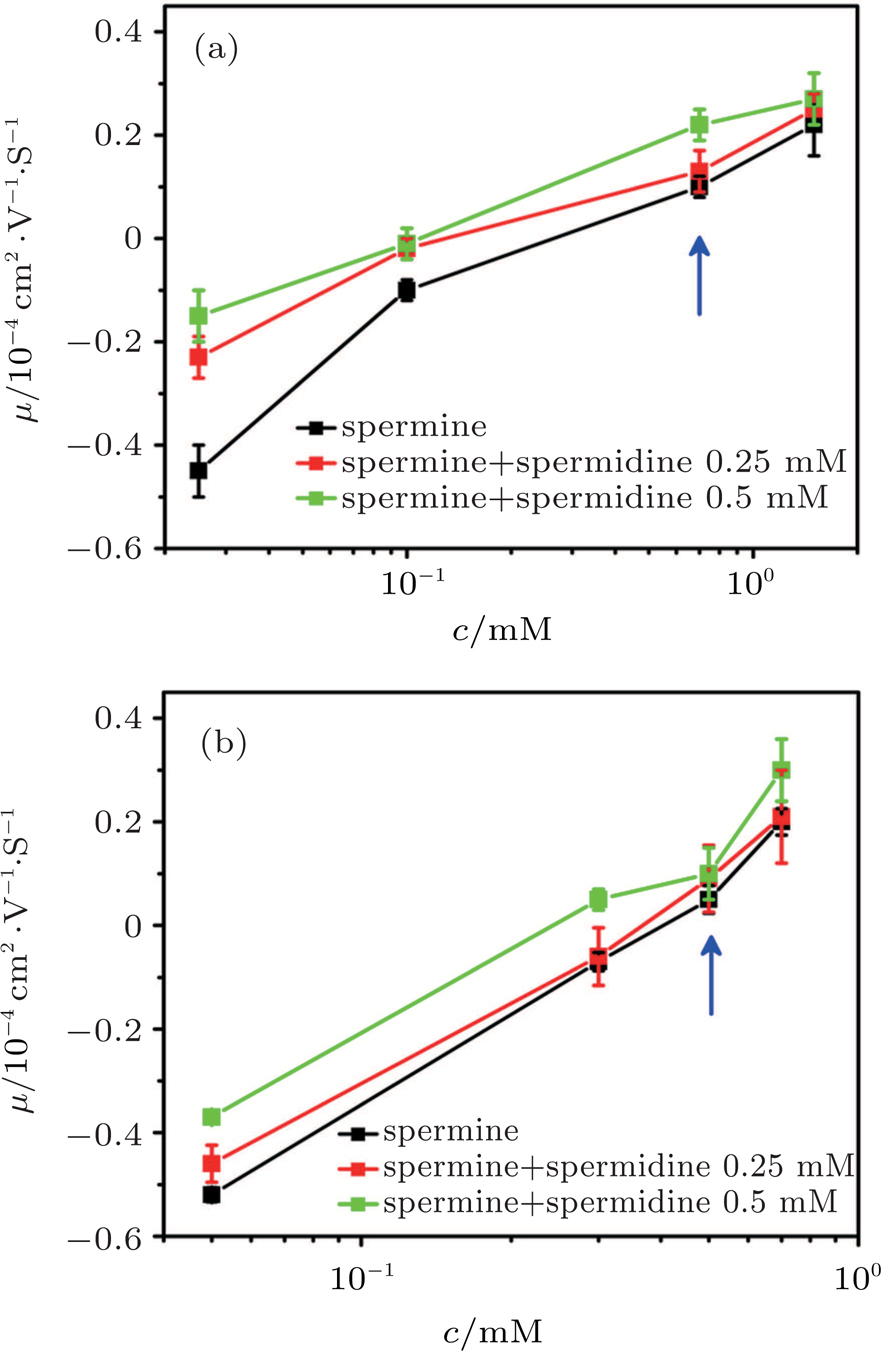 | Fig. 10. (color online) DNA molecule electrophoresis’ mobility μ as a function of the spermine concentration with trivalents permidine. (a) DLS measurements for 0, 0.25 mM, 0.5 mM of spermidine (black, red, green); (b) SME measurements for 0, 0.25 mM, 0.5 mM of spermidine (black, red, green). The arrows indicate increasing concentration of the spermidine. |
The suppression mechanism is proposed in Ref. [14]. They thought that the monovalent and divalent counterions were full of the strong correlation liquid to prevent the other ions entering inner DNA. They proposed an explanation for the re-entrant condensation based on a new theory of screening of macroions by multivanlent cations, emphasizing the strong corrections of multivalent cations at the surface of DNA.[37]
When more than one counterionis present in solution, the theory on the competitive electrostatic binding to polyelectrolytes is developed by Rouzina and Bloomfield in the framework of the nonlinear Poisson–Boltzmann equation.[20] Unfortunately, the theory cannot be applied to our system directly since there is no CI in PB theory. However, if the influence of strong correlation of counterions at the surface of a macrion is included as a boundary condition, the modified PB theory can elucidate the CI. For the current mixing couterions system, we can incorporate Rouzina and Bloomfield’s theory to SCL framework developed by Shklovskii group.[29,38] In the presence of competing cations of any valency, following the similar derivation of Ref. [20], we can have the general expression for the net linear charge density of DNA
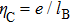

6. DNA compaction and charge inversion induced by organic monovalent ions
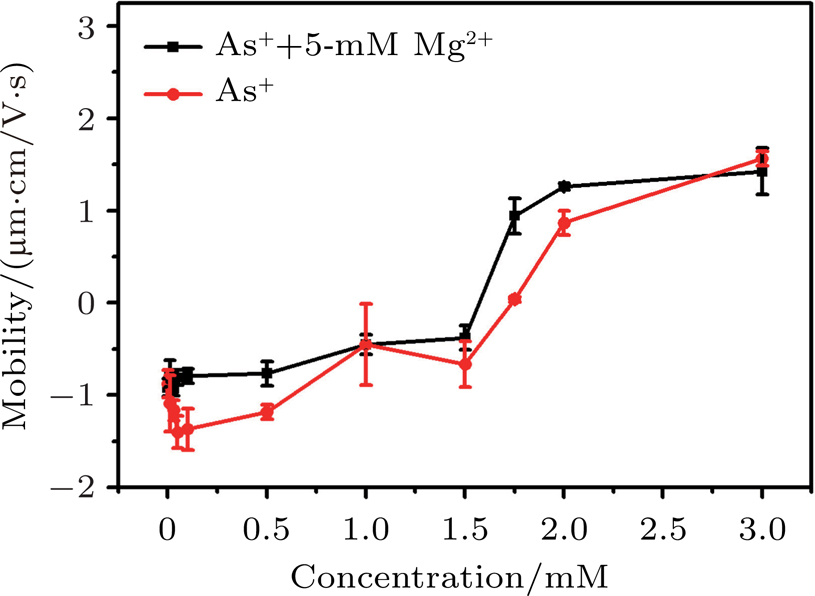
It has been proposed that electrostatic correlations are dominant and lead to CI in the case of counterions with valences larger than two near strongly-charged interfaces. In the case of monovalent ions, electrostatic correlations are weak and no CI is expected. However, when hydrophobic effect is introduced, scenarios may become quite different. Ph4As+ is an organic monovalent ion, and has strong hydrophobic effect.[39] When introducing the organic monovalent ion Ph4As+ into a DNA system, we can find that it not only provokes the CI of DNA, but also leads to DNA compaction. We measured the electrophoretic mobility of DNA in the mixture solution with different concentrations of Ph4As+ and 5-mM MgCl2 by DLS technique. The result is shown in Fig.
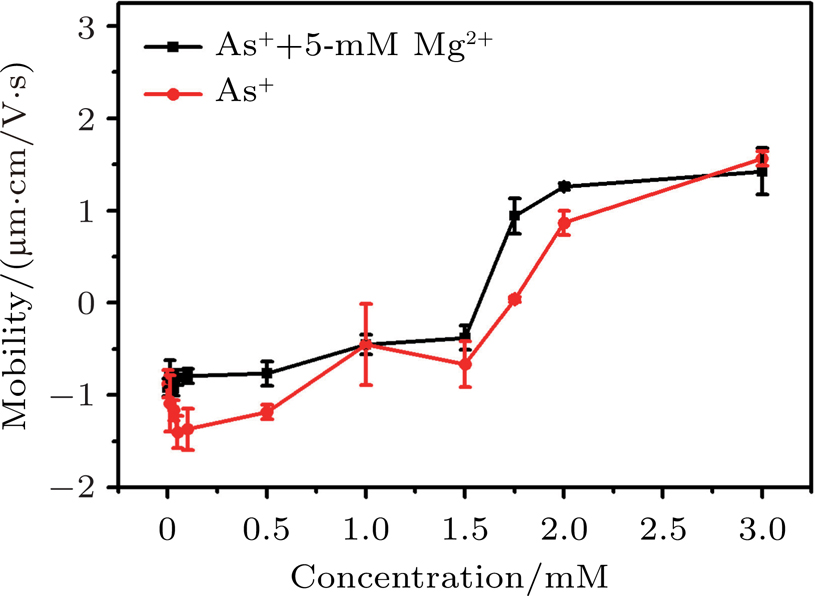 | Fig. 11. (color online) Electrophoretic mobility of DNA versus the concentration of Ph4As+. |
The mechanism of DNA charge reversal is similar to colloid CI, the hydrophobic effect as a driving force for CI. This mechanism is able to induce CI at low concentrations of monovalent ions containing hydrophobic functional groups, and saturation effects appearing at higher concentrations.
Following the similar analysis in Ref. [21], the critical concentration of counterions in solution when CI occurs can be expressed as

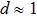
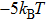
On the other hand, figure 
 | Fig. 12. (color online) AFM observation of DNA complexes. Panel (a) λ-DNA in 5-mM Mg2+ solution; Panels (b)–(f) DNA conformations at different concentrations (0.01, 0.05, 0.1, 0.5, and 1 mM, respectively) of As+ in 5-mM Mg2+ buffer. |
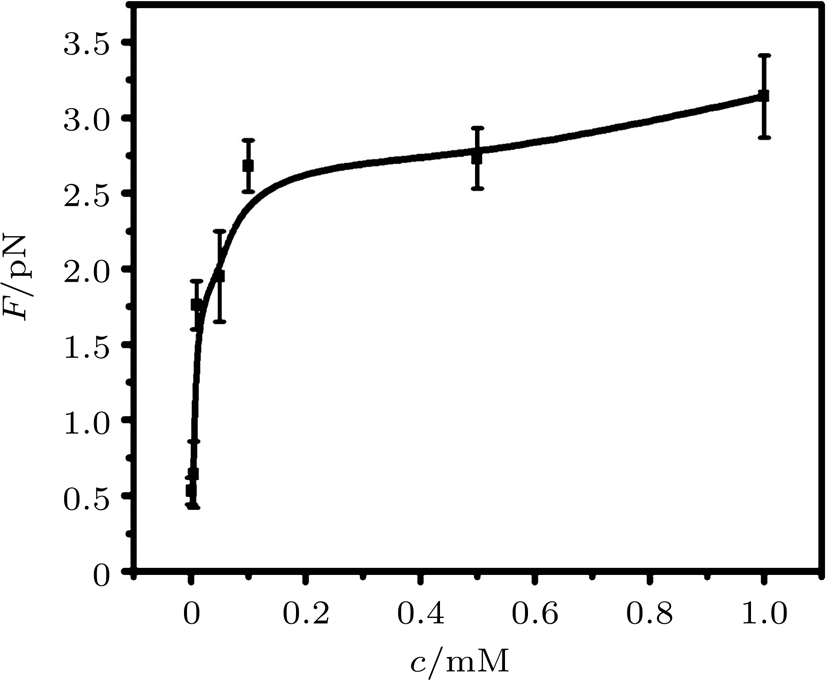
Figure
 | Fig. 13. The curve of condensing force at different concentrations of Ph4As+. |
7. Conclusion and perspectives
In this review, we presented the recent experimental progress on the modulation and control of DNA charge neutralization and inversion. The main results can be summarized as follows:
(i) The occurrence of DNA CI in trivalent counterion solution can be induced by adding ethanol to lower the dielectric constant of solution. In the meanwhile, we accomplished modulation of EM of DNA in two ways by decreasing or increasing dielectric constant of solution, infering that CI of DNA is a purely electrostatic phenomenon driven by the existence of a SCL of counterions at the DNA surface.
(ii) The charge neutralization and CI of DNA in couterion solution is dependent on pH value, which is related with the effective valance mechanism of couterion. Small and highly mobile hydrogen ions tend to attach DNA-counterion complex to further neutralize its remnant charge, which is related with the surface area of complex.
(iii) DNA Cl can be suppressed or promoted by mixing quadrivalent counterion (spermine) with low valance counterions. It was shown that the electrophoretic mobility of DNA in spermine solution decreases in the presence of monovalent sodium ions and divalent magnesium ions. This means that the charge neutralization of DNA by the quadrivalent counterions is suppressed when adding extra mono- or divalent counterions. The charge neutralization and inversion of DNA by quadrivalent counterions can be suppressed by mixing with mono- and divalent ions. In that case, the charge neutralization and inversion of DNA is promoted rather than suppressed by the additional trivalent ions.
(iv) Hydrophobic effect can be the main driving force of DNA CI and compaction. We show that the organic monovalent ions of tetraphenyl chloride arsenic (Ph4As+) can induce DNA compaction and even invert its electrophoretic mobility.
The synergy of DLS, electrophoretic mobility, single molecular force spectroscopy and different microscopy approaches allowed us an accurate characterization of the compaction and charge neutralization of the system. Although a quantitative comparison to theoretical models is difficult, due to the complexity of the system, our recent experimental results support the physical driving mechanism of DNA charge neutralization and compaction. Nevertheless, some other factors, such as preferential solvation and hydration might simultaneously play a role on CI of DNA complex. More experimental, theoretical, and numerical simulation efforts in a wider range of variation of different parameters (such as ionic strength, pH, counterion binding sites) are probably needed to shed light to the detail mechanism of DNA charge inversion and compaction.
Reference
| [1] | |
| [2] | |
| [3] | |
| [4] | |
| [5] | |
| [6] | |
| [7] | |
| [8] | |
| [9] | |
| [10] | |
| [11] | |
| [12] | |
| [13] | |
| [14] | |
| [15] | |
| [16] | |
| [17] | |
| [18] | |
| [19] | |
| [20] | |
| [21] | |
| [22] | |
| [23] | |
| [24] | |
| [25] | |
| [26] | |
| [27] | |
| [28] | |
| [29] | |
| [30] | |
| [31] | |
| [32] | |
| [33] | |
| [34] | |
| [35] | |
| [36] | |
| [37] | |
| [38] | |
| [39] | |
| [40] | |
| [41] |