


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Biexponential distribution of open times of a toy channel model
Cite this Article
Li Xiang, Zhong Jing-Jing, Gao Xue-Juan, Wu Yu-Ning, Shuai Jian-Wei, Qi Hong. Biexponential distribution of open times of a toy channel model. Chinese Physics B, 2017, 26(12): 128703 
Permissions
Biexponential distribution of open times of a toy channel model
† Corresponding author. E-mail:
Project supported by the National Natural Science Foundations of China (Grant Nos. 11504214, 31370830, and 11675134), the 111 Project, China (Grant No. B16029), and the China Postdoctoral Science Foundation (Grant No. 2016M602071).
Abstract
The biexponential distributions of open times are observed in various types of ion channels. In this paper, by discussing a simple channel model, we show that there are two different schemes to understand the biexponential distribution of open times. One scheme is mathematically strict based on generator matrix theory, while the other one has a clear physical explanation according to an approximation process with numerical simulation of Markovian channel dynamics. Our comparison results suggest that even for biologically complex channels, in addition to carrying out a stochastic simulation, the strict theoretical analysis should be considered to understand the multiple exponential distributions of open times.
1. Introduction
Ion channels play an important role in neuronal activities and intracellular signaling. The gating kinetics of ion channels has been extensively studied by analyzing the stochastic on/off flickering of single current traces obtained via patch clamp recording.[1] The most basic statistical parameters measured are the open probability 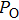
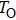
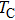


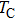
Typically ion channels are comprised of several subunits. For example, the voltage-dependent sodium (Na+) channel and potassium (K+) channel, the inositol 1,4,5-trisphosphate receptor (IP3R) calcium channel, and the ryanodine receptor (RyR) calcium channel are all tetramers. Different classes of gating models have been used to incorporate the structure of tetramer ion channels: models with independent and different subunits,[3] models with independent and identical subunits,[4–7] and allosteric models with cooperating subunits,[8,9] as well as the hybrid models.[10,11]
As an important tetramer channel, IP3R can release calcium ions (Ca2+) into cytosol from endoplasmic reticulum (ER) to modulate various intracellular functions. Single-channel properties of the Xenopus IP3R channel were examined by patch clamp electrophysiology of the outer nuclear membrane of isolated oocyte nuclei, giving a biexponential distribution of open-channel dwell times with time constants of about 4 ms and 20 ms, respectively.[12] The biexponential distribution of open time has also been observed in other channels, including the Ca2+ channels in the A7r5 smooth muscle-derived cell in the presence of dihydropyridine agonists,[13] the RyR Ca2+ channel of skeletal muscle in the presence of bastadin 5 and bastadin 10.[14,15]
An interesting question is the dynamics for biexponential distribution of open times of channels. The simple consideration is to directly assume two open states which determine the two exponential distributions, i.e., a short open state responding to a fast decay distribution of open times and a long open state responding to a slow decay distribution. Following this scheme, a channel model with five open states has been considered to exhibit the biexponential distribution of open times for the Ca2+ channels in the A7r5 smooth muscle-derived cell in the presence of dihydropyridine agonists.[13] A different scheme for biexponential distribution of open times has been proposed in Ref. [16]. Such a channel model consists of four identical, independent subunits, each of which has only one active state. The channel opens when there are at least three subunits in the active state. The model successfully reproduces the biexponential distribution of open times.[16,17]
In Refs. [16] and [17], the IP3R channel consists of four identical and independent subunits, each with the 9-state. In order to discuss clearly the origin of the behavior of biexponential distribution of open times, a toy model is considered in this paper. According to the theoretical analysis and numerical simulation of the toy model, we indicate that there are two different schemes to understand the biexponential distribution. One scheme has a clear physical explanation based on an approximation process, but a better scheme is mathematically strict with little physical picture.
2. Toy channel model
In the paper, we consider a toy channel model. The channel is composed of two identical and independent subunits. Each subunit has only two states: the active state (O) and the rest state (C):
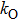
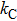
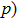
For the channel model, there are three states, i.e., CC, CO, and OO states. We assume that once there is a subunit in active state, the channel is defined as being open. So the channel states OO and CO are open states, while the channel state CC is a closed state. Accordingly, the three channel states have the following transition processes:
To consider dimensionless unit by assuming 
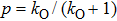
Thus in the model, we have only one free parameter p which is the active probability of the subunit. So the channel model can be rewritten as
As a result, the channel open probability 


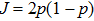
3. Theoretical analysis of open-time distribution
In the following, we use the generator matrix theory[16,18,19] to discuss the open-time distribution of the channel. Assuming that the probabilities in the states of CC, CO and OO are C, O1, and O2, respectively, the binding-unbinding reaction equations of the channel are given by
Considering the definition of the exponent of a matrix,


Let
As a result, the expression 



One property of the open-time distribution is that the total open-time distribution should be unity. In order to check it, we consider the integral as follows:

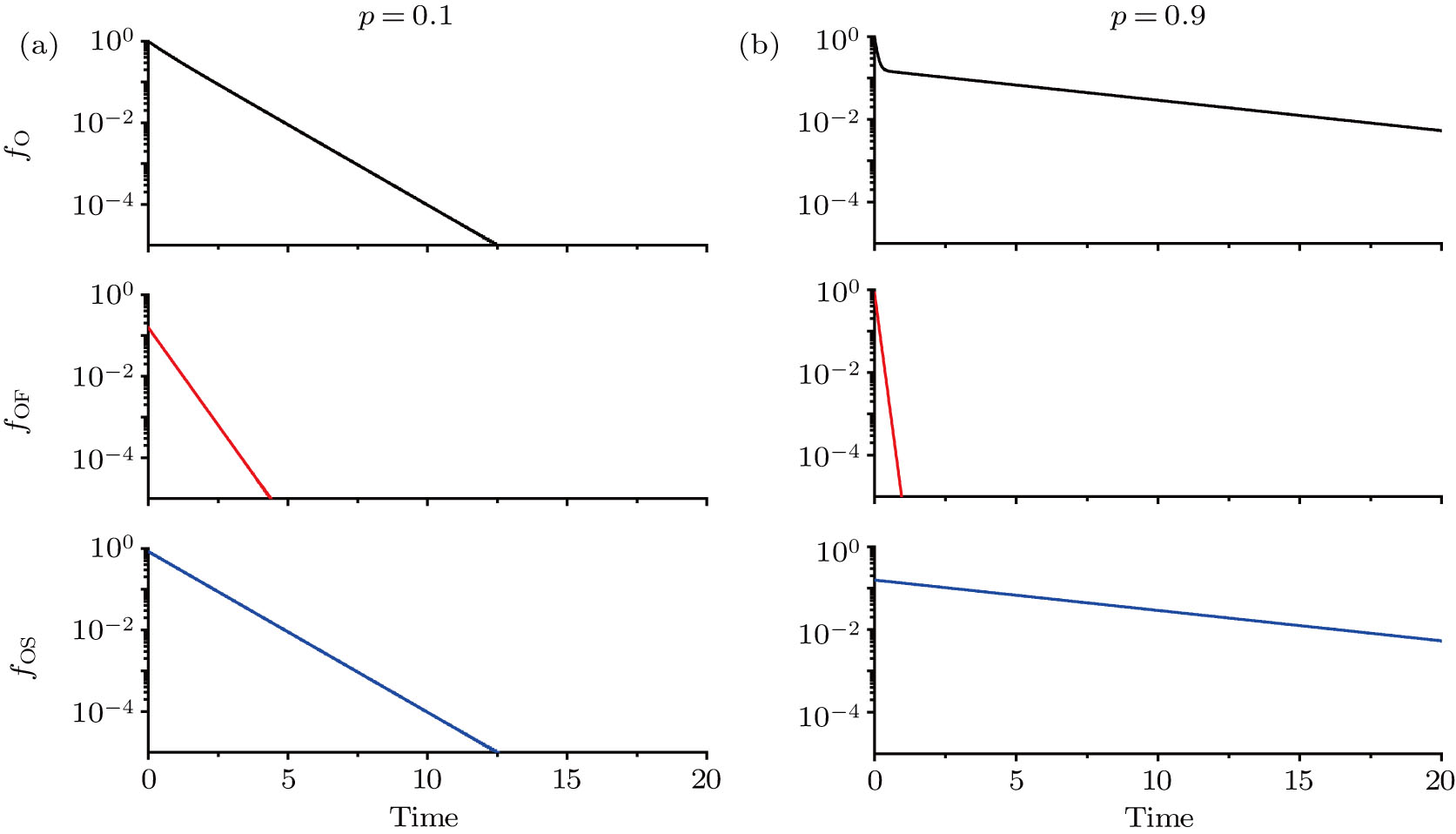
Figure 

 | Fig. 1. (color online) Open-time distributions at p = 0.1 (a) and 0.9 (b). The upper, middle and lower panels give the total, the fast decay, and the slow decay distributions of open times, respectively. In the model, the time has a dimensionless unit. |
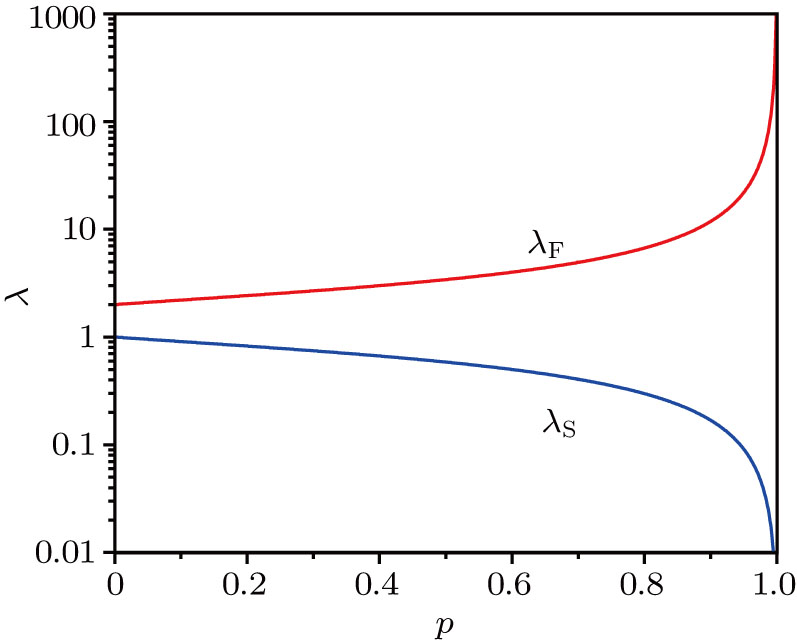
Figure 


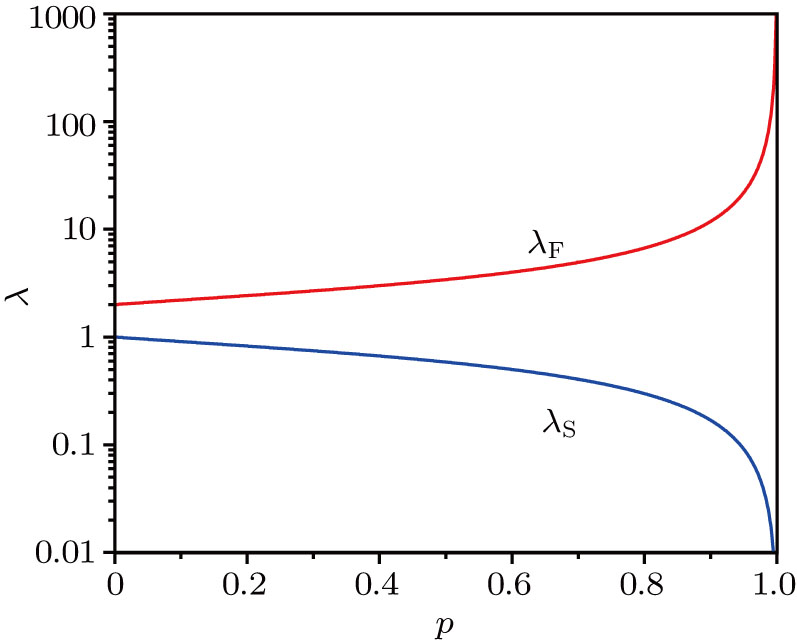 | Fig. 2. (color online) Two exponential decay rates versus p, obtained in theory. |
Figure
 | Fig. 3. (color online) Contribution percentages of (a) fast decay mode and (b) slow decay mode to the open-time distribution. |
4. Markovian simulation of open-time distribution
Now we carry out the numerical simulation to discuss the open-time distribution of the channel model. As a result, the stochastic simulation of the detailed transition process between active and rest states of each subunit is recorded to obtain the stochastic time series of channel open and closed states. Different approaches have been suggested to simulate the stochastic channel dynamics.[20–22] In the paper, we directly simulate the stochastic dynamics of the channel model by a two-state Markovian process.[5,23,24] In detail, the state of each subunit is updated in small time steps of 


In the following, by discussing the Markovian numerical simulation, we provide another scheme to understand the biexponential distribution of open times. Figure
 | Fig. 4. (color online) Variations of (a) channel state and (b) the number of active subunits with time at p = 0.5. The channel open state can be classified as the CO-only and OO-related open states as marked by arrows. |
With the trajectories of channel open and close states, we can calculate the open time distribution. As examples, the open-time distributions at p = 0.1, 0.5, and 0.9 obtained with Markovian simulation are plotted in Fig.
 | Fig. 5. Open-time distributions at p = 0.1 (a), 0.5 (b), and 0.9 (c), obtained with Markovian simulation. |
Because the channel open states can be distinguished as the CO-only and OO-related open states, the open time series can then be split into the CO-only and OO-related open modes. The distributions of the CO-only and OO-related open time events are given in Figs.
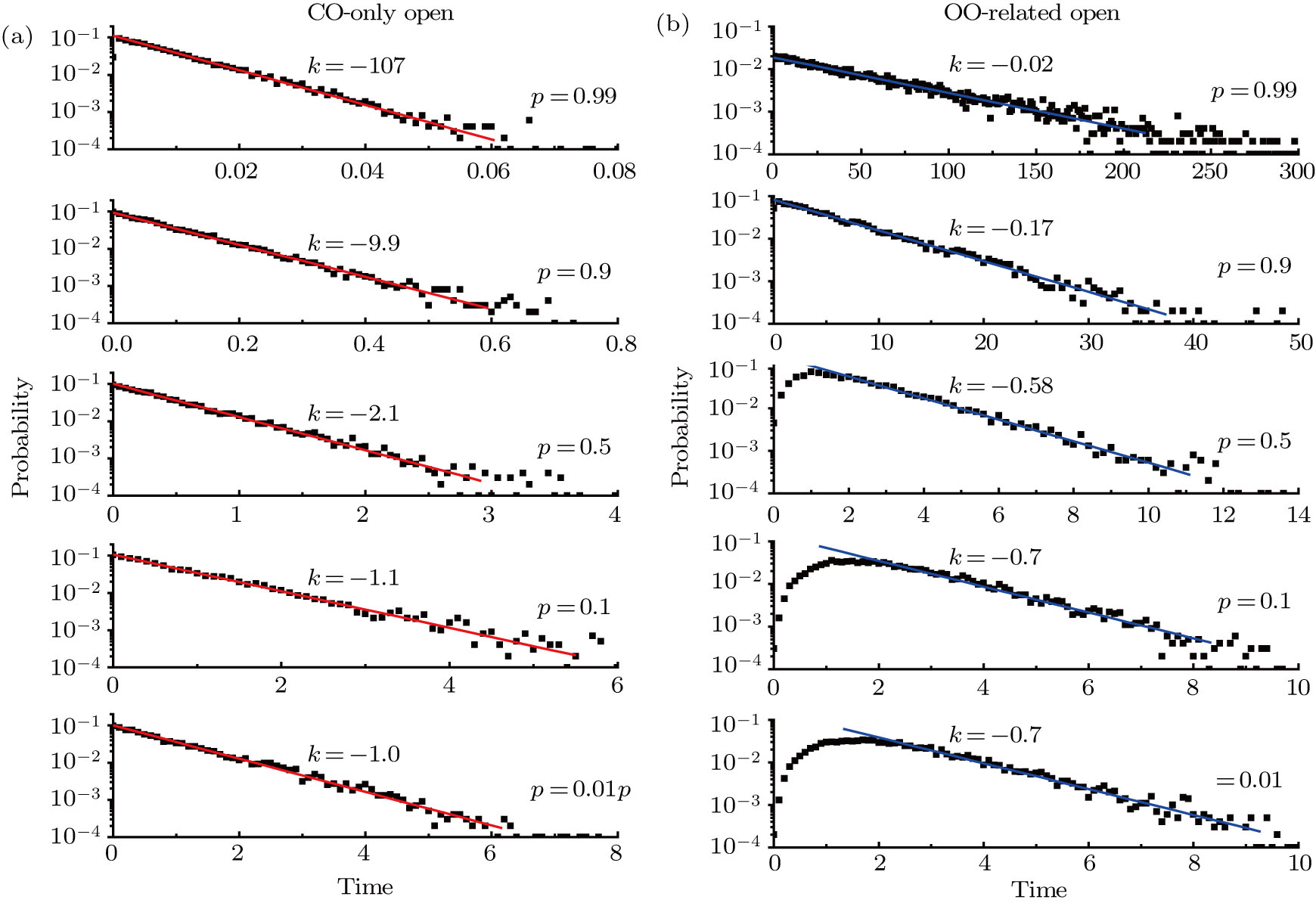
 | Fig. 6. (color online) Distributions of (a) CO-only open events and (b) OO-related open events at p = 0.99, 0.9, 0.5, 0.1, and 0.01 from upper to lower panels. The values of decay rate k fitted with the exponential function are also given in each sub-figure. |
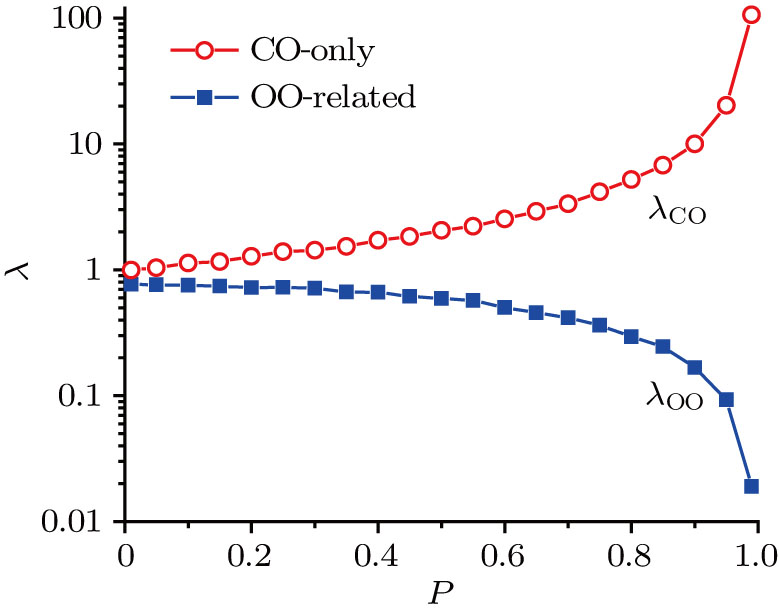
By fitting the exponential decays of CO-only and OO-related open-time distributions, the exponential decay rates can be obtained. The decay rates for both the CO-only and OO-related open states against p are given in Fig.
 | Fig. 7. (color online) Exponential decay rates of the CO-only and OO-related open-time distribution versus p. |
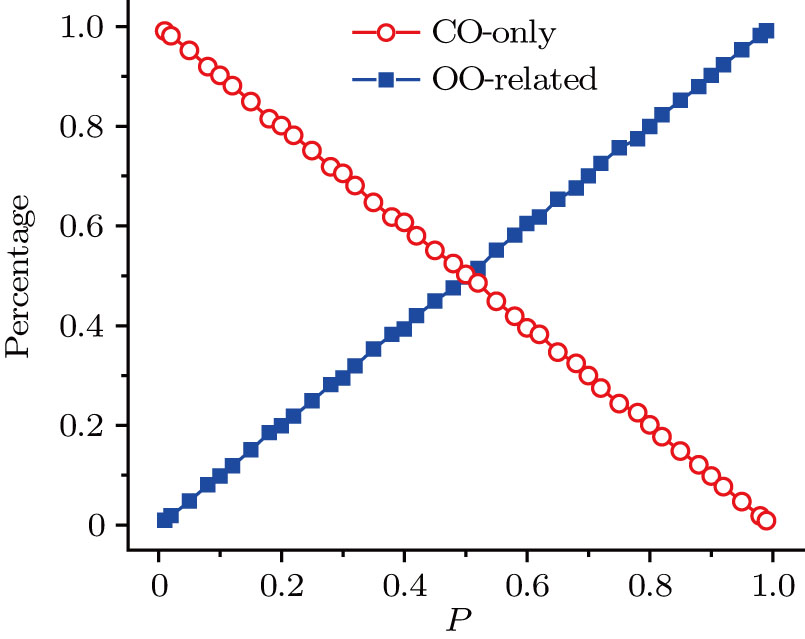
By counting the numbers of CO-only and OO-related open events of the open time series, the contribution percentages of CO-only and OO-related open modes as a function of p can be calculated as given in Fig.
 | Fig. 8. (color online) Plots of contribution percentage of CO-only and OO-related open modes versus p. |
5. Conclusions and perspectives
In this paper, we consider a toy model to discuss the behavior of the biexponential distribution of open times, which has been observed in various types of channels.[12–15] The toy channel consists of two identical and independent subunits. Each subunit has only an active state and a rest state. The channel opens when at least one subunit is in active state. The open time distribution of the channel is investigated systematically with theoretical analysis and Markovian simulation. We show that there are two different schemes to understand the biexponential distribution of open times.
First we use the generator matrix theory[16,18,19] to discuss the open-time distribution. Mathematically strict results are derived, but with little physical picture to understand clearly what open time events are fast and slow decay modes. The biexponential distribution of open times actually originates from 2 by 2 square open matrix, due to the two open states of CO and CC. As a result, the expression of open-time distribution consists of two exponential decay modes: one with a slow decay rate and the other with a fast decay rate. A surprising observation is that the fast and slow decay modes contribute to the open time series in a nonlinear manner with increasing p. The largest percentage that the fast decay contributes to the open time series at p = 0.5 is still less than 15%.
Another scheme on open-time distribution has a clear physical explanation, but based on an approximation process for the fitting of exponential distribution. Because the channel has two open states, i.e., CO-only and OO-related open states, the open time events can then be split into the CO-only and OO-related open modes. Linearly increasing and decreasing curves are observed for contribution percentages of OO-related and CO-only open modes, respectively. Especially, at p = 0.5, the CO-only and OO-related open states each contribute 50% probability to the open time events.
As a result, the two different schemes reveal different characteristics of the open times of the channel model. The scheme with the classification of CO-only and OO-related open states is easily understandable with a clear physical picture, which has been used to discuss the behavior of the open time distribution of the IP3R channel model based on the numerical simulation of stochastic channel dynamics.[16,17] Compared with the toy model discussed here, the biologically realistic IP3R channel model is complex. Thus it is a little hard to use the generator matrix theory to investigate the open-time distribution of the IP3R channel model. However, in the present paper our comparison results suggest that the analytic discussion with the strict generator matrix theory should be considered in order to understand the behaviors of multiple exponential distributions of open times of biological channels, such as the IP3R channel, which are typically complex.
Reference
| [1] | |
| [2] | |
| [3] | |
| [4] | |
| [5] | |
| [6] | |
| [7] | |
| [8] | |
| [9] | |
| [10] | |
| [11] | |
| [12] | |
| [13] | |
| [14] | |
| [15] | |
| [16] | |
| [17] | |
| [18] | |
| [19] | |
| [20] | |
| [21] | |
| [22] | |
| [23] | |
| [24] |