{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Computational analysis of the roles of biochemical reactions in anomalous diffusion dynamics
[Rueangkham Naruemon1, Modchang Charin1, 2, 3, †,  ]
]
]
|
|


† Corresponding author. E-mail:
Project supported by the Thailand Research Fund and Mahidol University (Grant No. TRG5880157), the Thailand Center of Excellence in Physics (ThEP), CHE, Thailand, and the Development Promotion of Science and Technology.
Most biochemical processes in cells are usually modeled by reaction–diffusion (RD) equations. In these RD models, the diffusive process is assumed to be Gaussian. However, a growing number of studies have noted that intracellular diffusion is anomalous at some or all times, which may result from a crowded environment and chemical kinetics. This work aims to computationally study the effects of chemical reactions on the diffusive dynamics of RD systems by using both stochastic and deterministic algorithms. Numerical method to estimate the mean-square displacement (MSD) from a deterministic algorithm is also investigated. Our computational results show that anomalous diffusion can be solely due to chemical reactions. The chemical reactions alone can cause anomalous sub-diffusion in the RD system at some or all times. The time-dependent anomalous diffusion exponent is found to depend on many parameters, including chemical reaction rates, reaction orders, and chemical concentrations.
Diffusion is likely to be the most important transportation process inside a living cell. However, the cell cytoplasm is not a simple solvent; it is crowded and consists of many chemically reactive molecules. Therefore, various factors can hinder diffusion in living cells.[1,2] In particular, the chemical reactions or biochemical conversions in metabolic and signal transduction pathways can affect the cellular transportation processes.[1,3] In general, the reaction diffusion processes in a living cell can be described by a set of reaction–diffusion equations,
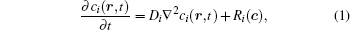
Anomalous diffusion is characterized by the MSD of diffusing particles, which grows with the power law of time:
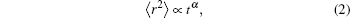
The ability of chemical reactions to cause anomalous sub-diffusion has not yet been experimentally investigated. However, it has been observed and clearly identified in many other cases. Many researches demonstrated that the continuous time random walk (CTRW) model is one of the most successful theoretical models of anomalous diffusion.[13,17,18] Also theoretical models based on the CTRW and fractional reaction–diffusion equations confirmed that chemical components can cause anomalous sub-diffusion.[13,17,19–23] Likewise, Haugh used the framework of Green’s function analysis to demonstrate that the reaction diffusion mechanism could not be assumed to be normal diffusion.[3] Using Monte Carlo simulations on a simple lattice model, Saxton also showed that the binding mechanism alone can cause anomalous diffusion at some or all times.[24] In addition, by using lattice gas automata, researchers have recently found that the diffusion in the Michaelis-Menten mechanism exhibits anomalous diffusion in a transient period.[25,26]
In this work, we further investigate the diffusive anomaly of a reaction diffusion system by using MCell, a 3D spatially realistic Monte Carlo simulator that tracks individual molecules.[27] Although the MCell program has been optimized for Monte Carlo simulation, a deterministic approach is computationally more efficient and easier to implement for most problems. Thus, we propose alternative methods using a deterministic algorithm to investigate the anomalous behaviors in reaction diffusion systems which have been numerously elucidated by stochastic simulation in published versions. The effects of reaction rate and reaction type on the diffusion behavior are also investigated.
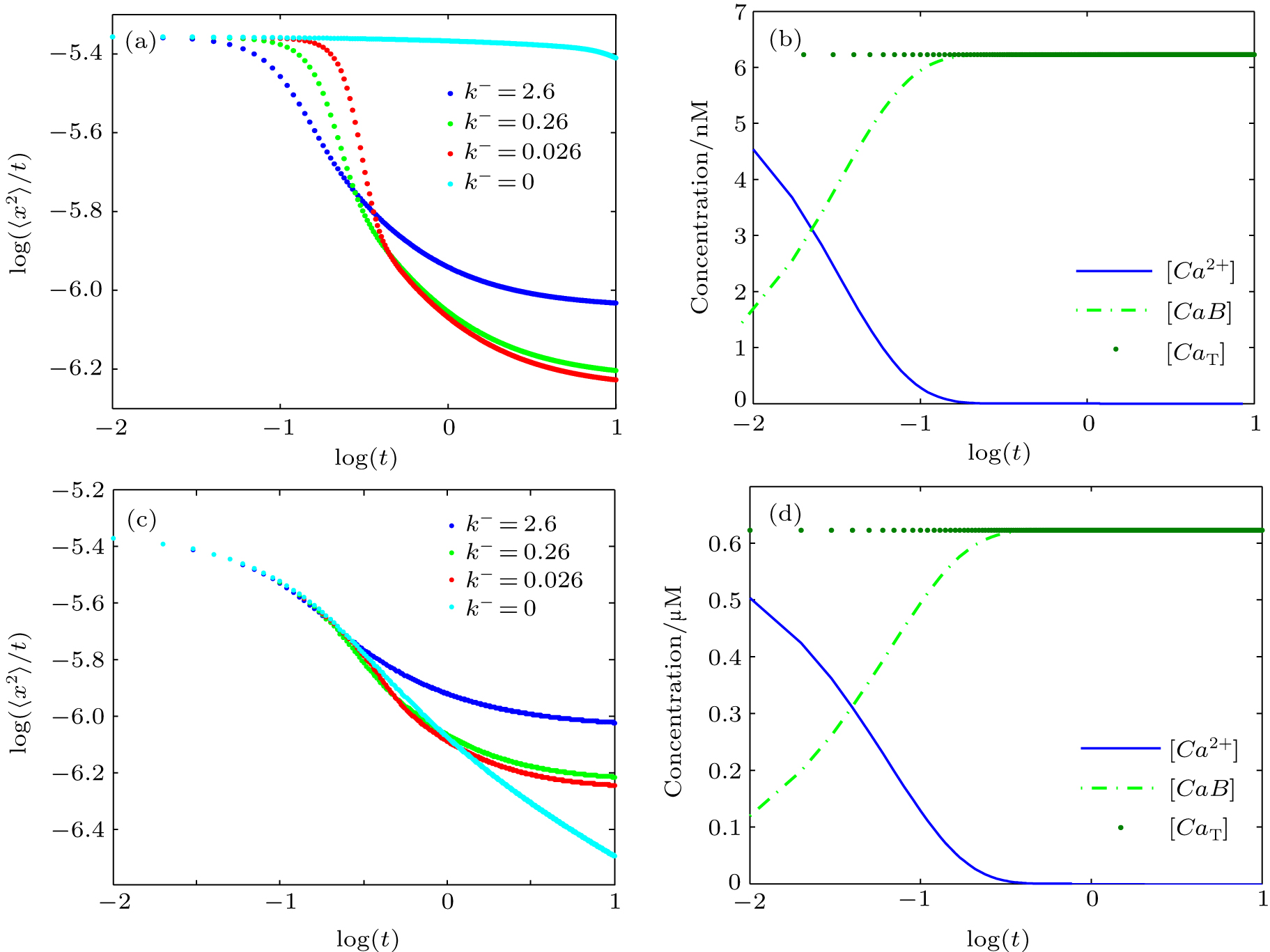
Reaction–diffusion equations are a mathematical model that explains the changes of one or more substances in space and time in chemical reaction and diffusion processes. In addition to the study of chemistry, these equations can also be used to describe many intracellular processes.[28,29] In the present work, we study a Ca2+ buffering system in cells. This Ca2+ buffering system can control various processes, including neurotransmitter release during synaptic transmission. The calcium buffering dynamics can be written in the form of a chemical equation as follows:
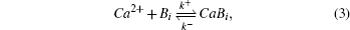
We investigate anomalous diffusion in the Ca2+ buffering system by using MCell version 3.1, a Monte Carlo (MC) algorithm that tracks individual molecules.[27] The MCell simulations are performed in three-dimensional (3D) space, and all of the Ca2+ trajectories are recorded during the simulations. The recorded trajectories are then used to calculate the mean squared displacement (MSD) of Ca2+. In all simulations, the dimensions of the computational geometry are 4 μm × 4 μm × 4 μm. Initially, 54902 ions of Ca2+ are held in a spherical shell with a radius of 7.5 nm located at the origin and are then released at time t = 0, whereas 549020 ions of buffers are uniformly distributed throughout the computational box. These numbers of ions correspond to a calcium concentration of 1.425 μM and a buffer concentration of 14.25 μM in our computational domain. All surfaces of the computational box are reflective to all molecules. To study only the reaction diffusion dynamics and avoid the possible effects of the boundary on calcium ion dynamics, we ensure that calcium ions and calcium bound buffers do not reach the boundaries of our computational box. All parameter values used in MCell simulations are adjusted to reduce the simulation time, as given in Table
Deterministic simulation is an alternative approach to the study of the diffusive behaviors in the system in order to obtain longer time courses of the MSD. Assuming mass action kinetics and Fickian diffusion, the concentration changes in Eq. (
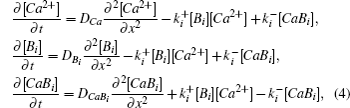
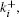
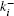
| Table 1. Parameter values in the simulated system. . |
The deterministic calculations that solve the differential equations in Eq. (
For the integration method, the MSD (〈x2〉) can be directly calculated from definition,[32]
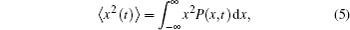
Different types of reactions are investigated to systematically study the effects of reactions in reaction diffusion systems.
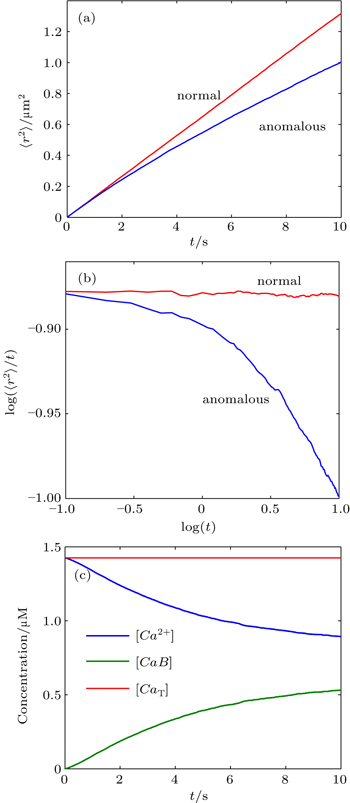
First, we used MCell, a particle-based Monte Carlo algorithm, to investigate anomalous diffusion in reversible bimolecular reaction, which can be found in most living cells. The particle trajectories in three-dimensional space are recorded and used to calculate their mean squared displacement (MSD or 〈r2〉) as shown in Figs.
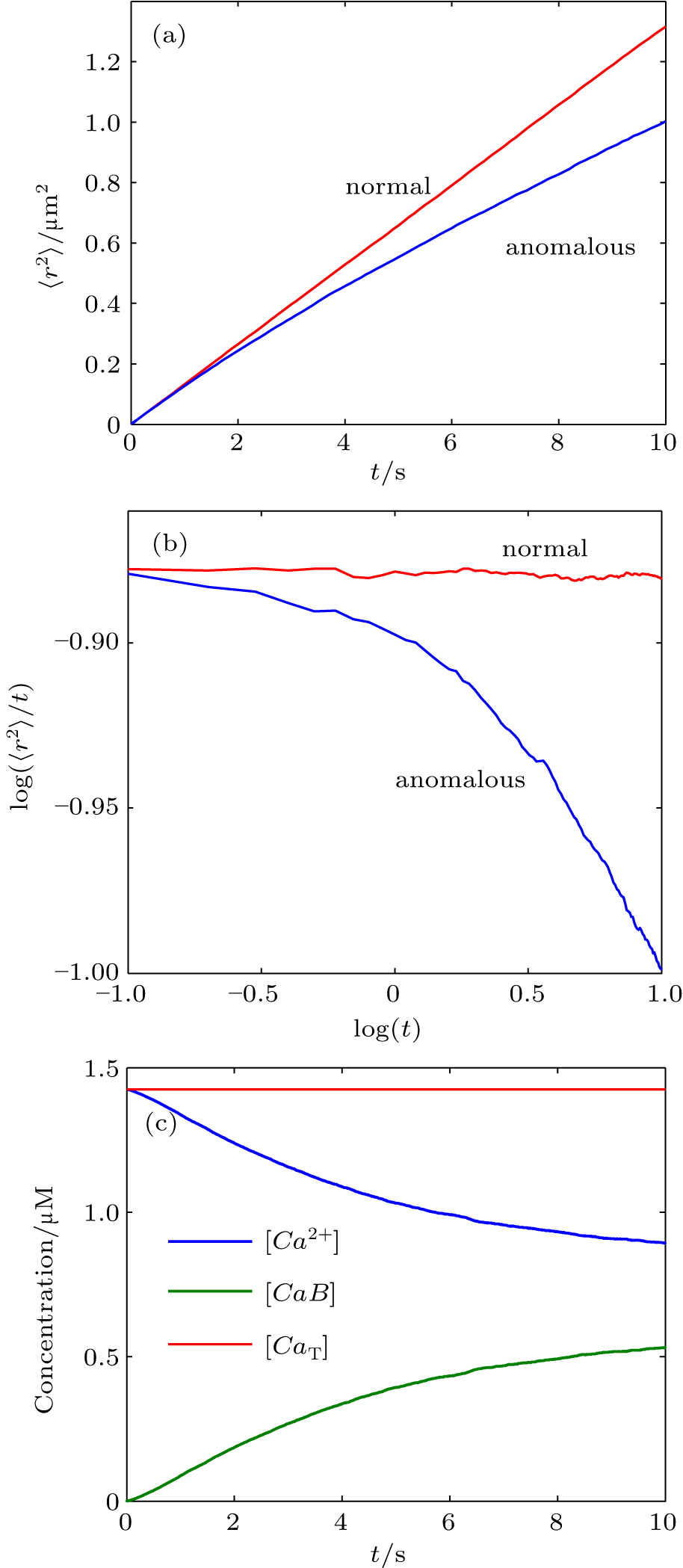 | Fig. 1. (a) Monte Carlo results for the mean-square displacement, 〈r2〉, as a function of time, t, for free diffusion (normal) and reaction diffusion (anomalous) in a linear plot. (b) Monte Carlo MSD is plotted as log (〈r2〉/t) versus logt. The slope of a straight line in this plot is α − 1; thus, normal diffusion dynamics yields a horizontal line. (c) The concentrations of the reactant, (Ca2+), and product, (CaB), as a function of time. The total calcium concentration, (CaT), is also shown. |
We cannot track the movement of individual molecules in the deterministic simulation; thus, the MSD cannot be directly computed from particle trajectories, and an alternative method to compute the MSD becomes necessary. We propose two alternative approaches to extract the MSD from the PDFs. In the deterministic calculations, we obtain PDFs at different times (P(x,t)) by solving the differential equations in Eq. (
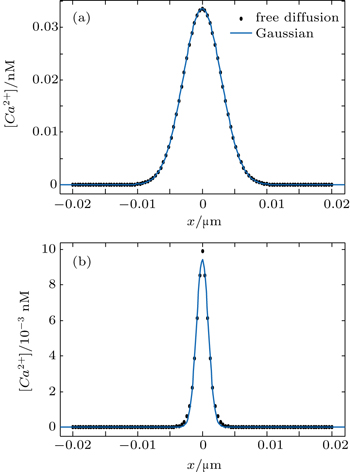
For the free diffusion in an infinite system with an initial [Ca2+] given by the Dirac delta function, the PDF can be written in the form of a Gaussian function[34] as verified in Fig. 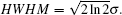
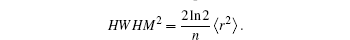
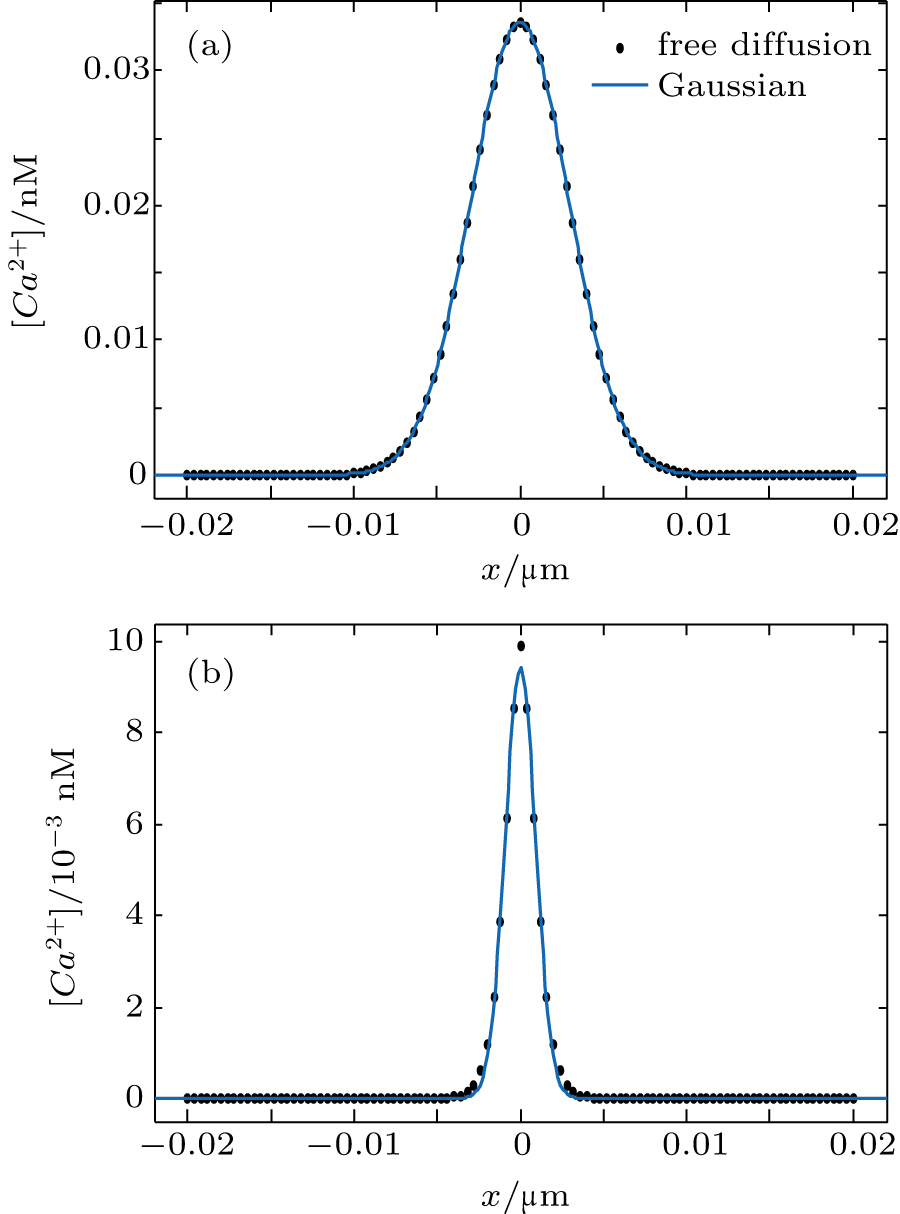 | Fig. 2. The [Ca2+] at t = 2 s (dots) are fitted to a Gaussian function (solid curves) for (a) free diffusion with the goodness of fit r-squared = 1 and (b) reaction diffusion with the goodness of fit r-squared = 0.997. |
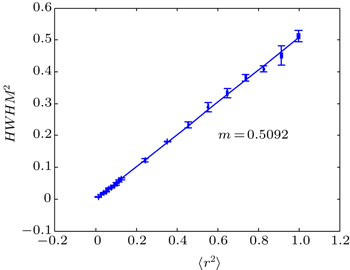
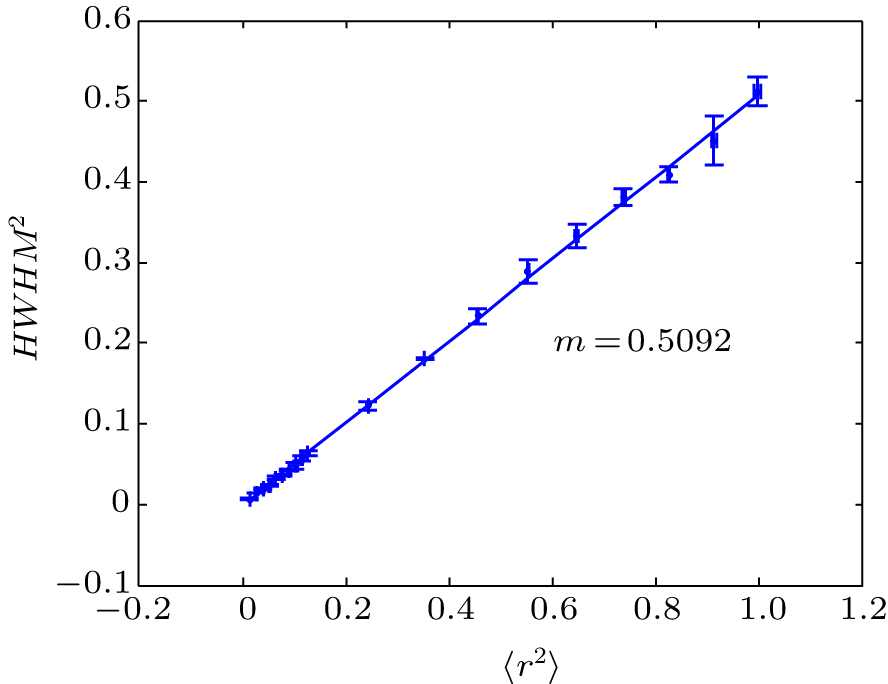 | Fig. 3. Relationship between HWHM2 and MSD (〈r2〉) in the reaction diffusion system. Simulation conditions and parameters are the same as those in Fig. |
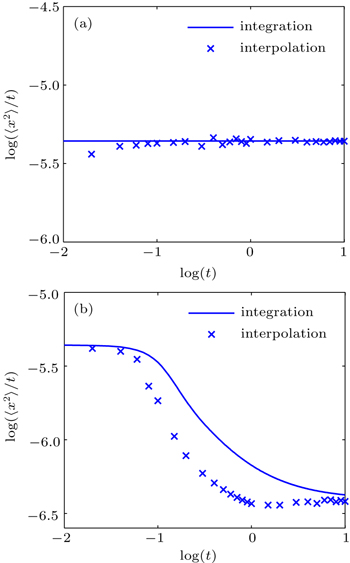
The MSD values obtained from HWHM interpolation and numerical integration for normal and anomalous diffusion are compared in Figs.
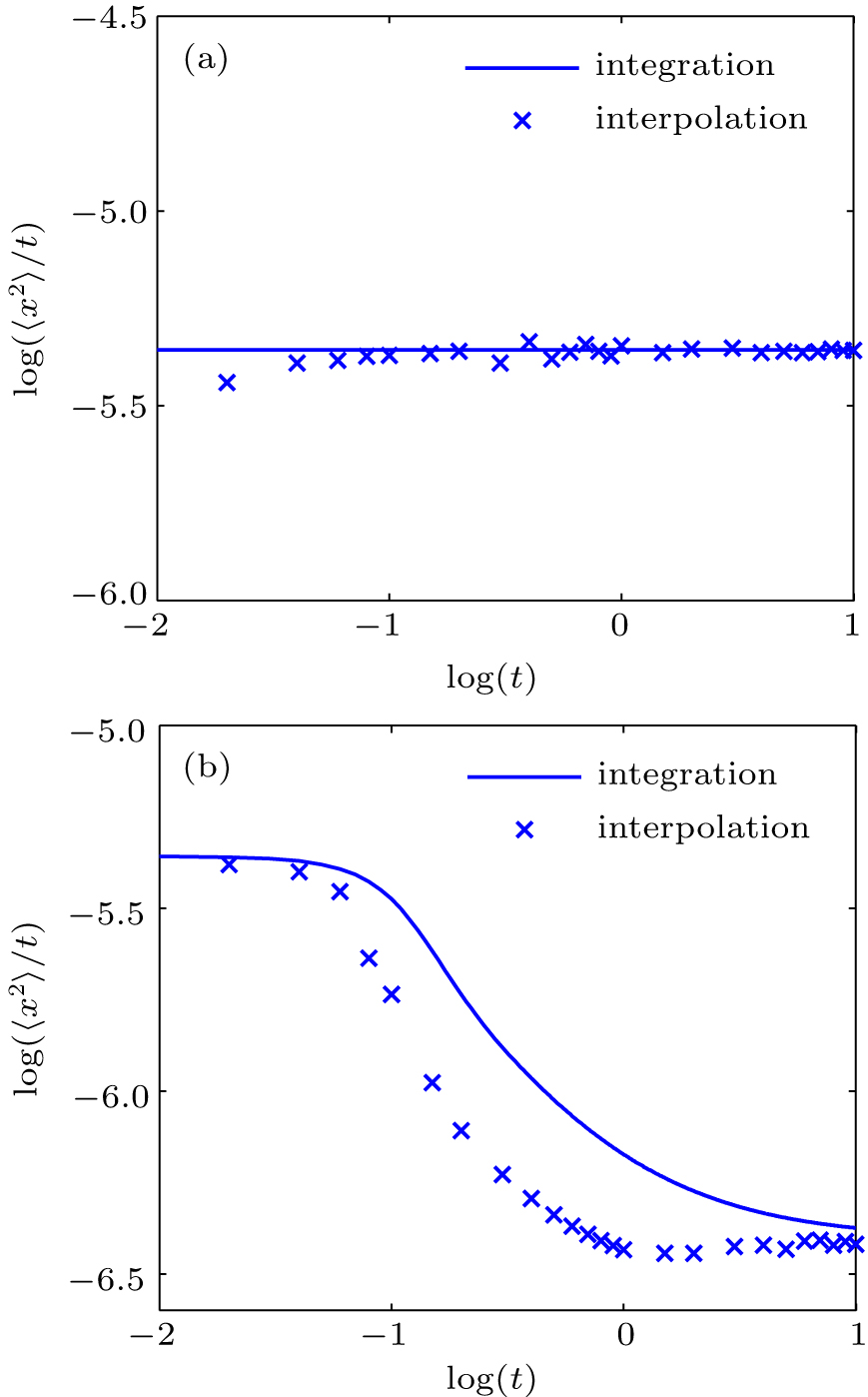 | Fig. 4. Comparisons between the MSD obtained from HWHM interpolation and that from numerical integration method for (a) normal diffusion and (b) anomalous diffusion. |
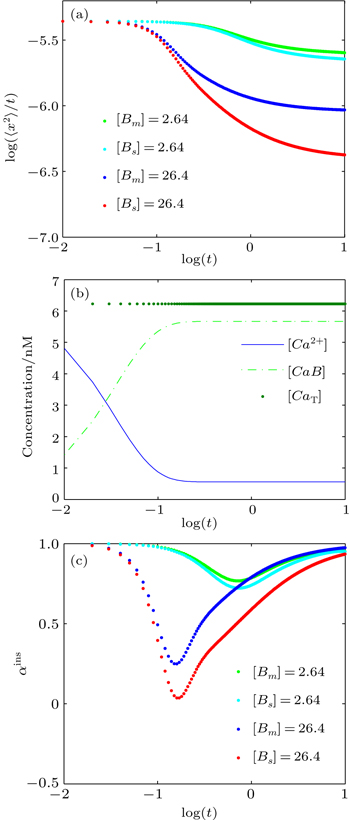
The reaction diffusion dynamics is henceforth investigated in one-dimensional space by using deterministic simulations in MATLAB. To study the anomalous behaviors of the system, the anomalous diffusion exponent (α) in the equation 〈x2〉 = Γtα is computed. As described above, the numerical integration method is selected to investigate the dynamics of reaction diffusion mechanisms and analyze the anomalous diffusion exponents (α). In this section, we focus on the reversible reaction of the Ca2+ buffering system. We investigate the effects of buffer concentration and diffusion coefficient on the diffusive behavior of Ca2+ in a buffer solution; the parameter values for this system are shown in Table
Figure
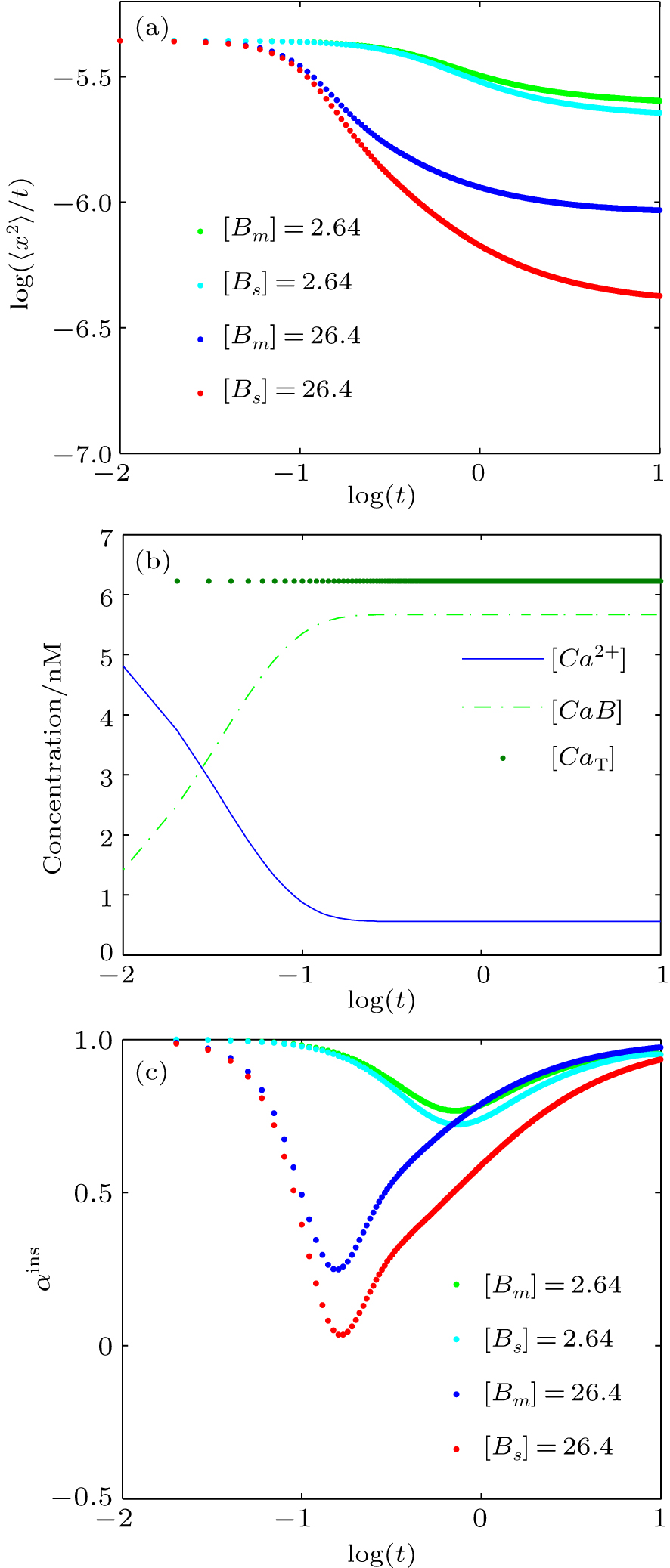 | Fig. 5. (a) Plots of log (〈x2〉/t) versus logt for Ca2+ in the presence of either stationary or mobile buffer at [Bi]0 = 2.64 μM and 26.4 μM; (b) time evolutions of Ca2+ and Ca2+-bound buffers (CaB) concentrations, where [Bm]0 = 26.4 μM, and the total calcium concentration (CaT) is also shown; (c) plots of the instantaneous anomalous diffusion exponent (αins) versus logt, for different values of [Bm] and [Bs]. |
Because the MSD can be expressed as
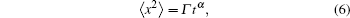
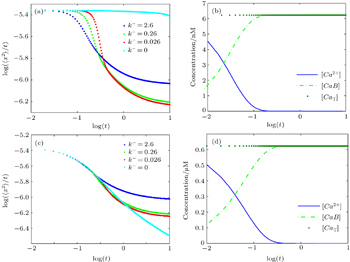
Figure 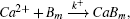
 | Fig. 6. (a) Plots of log (〈x2〉/t) versus logt for various values of dissociation rate (k−) in the case of calcium diffusion in mobile buffers at [Bm]0 = 26.4 μM and [Ca2+]0 = 6.23 nM; (b) concentration changes of Ca2+, CaBm, and CaT for k− = 0; (c) plots of log(〈x2〉/t) versus logt for various values of dissociation rates (k−) in the case of calcium diffusion in mobile buffers at [Bm]0 = 26.4 μM and [Ca2+]0 = 0.623 μM; (d) time-dependent concentration variations of Ca2+,CaBm, and CaT for k− = 0. |
To demonstrate the effect of the initial Ca2+ concentration, we also change [Ca2+]0 to 0.623 μM. The results in Fig.
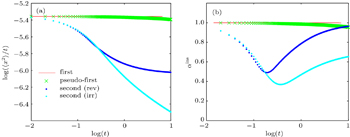
For the first-order reaction 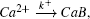
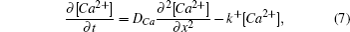
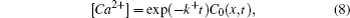
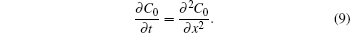
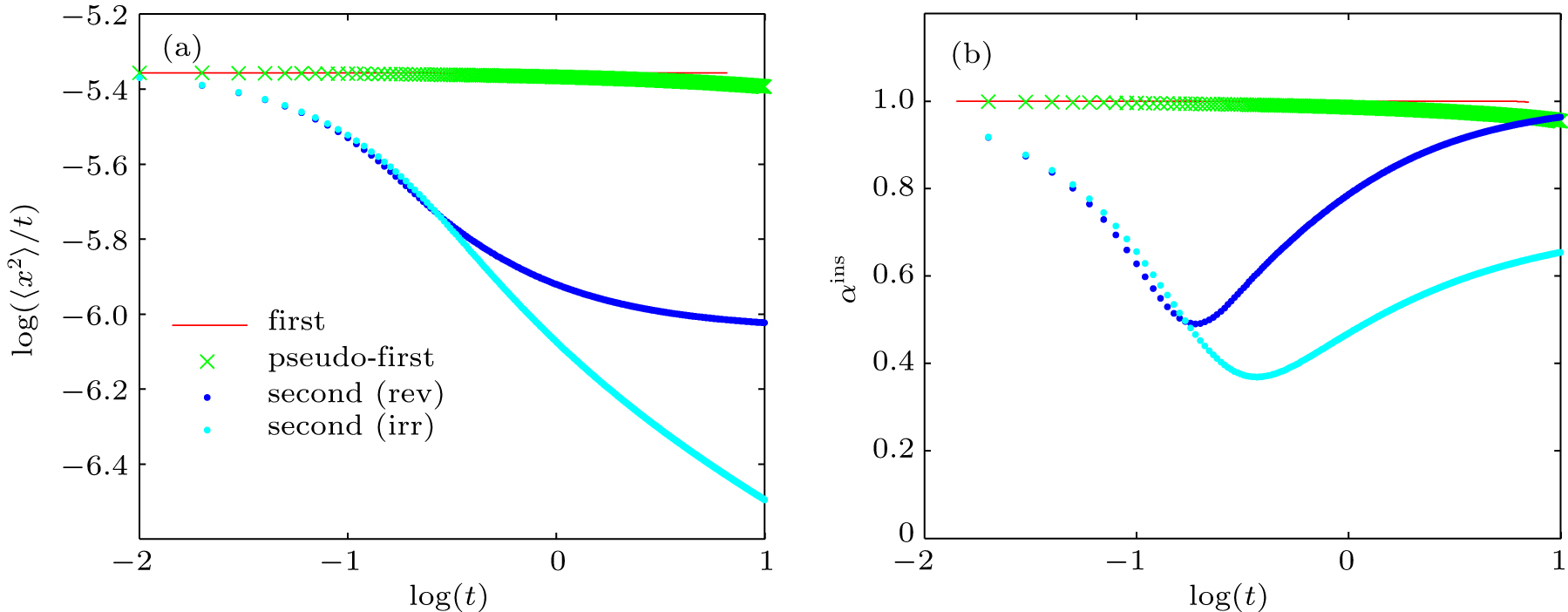 | Fig. 7. Time evolutions of (a) MSD and (b) instantaneous anomalous diffusion exponents (αins) for different types of reactions. |
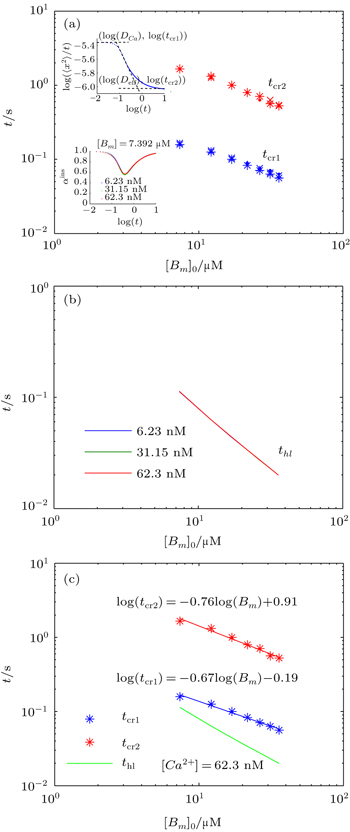
To calculate the average anomalous diffusion exponent (αavr) in the period of anomalous sub-diffusion, we first find crossovers between normal and anomalous diffusion by plotting log(〈x2〉/t) versus log(t). As shown in previous sections, in the case of a reversible second-order reaction, there are two major transitions at short time and at long time respectively. At the first crossover (x1,y1) diffusion changes from normal to sub-diffusion and at the second crossover (x2,y2) diffusion changes from sub-diffusion to normal diffusion again. To find these crossovers, we locate the intersections between two straight lines in anomalous and normal regions at early time and at long time as shown in the upper inset of Fig.
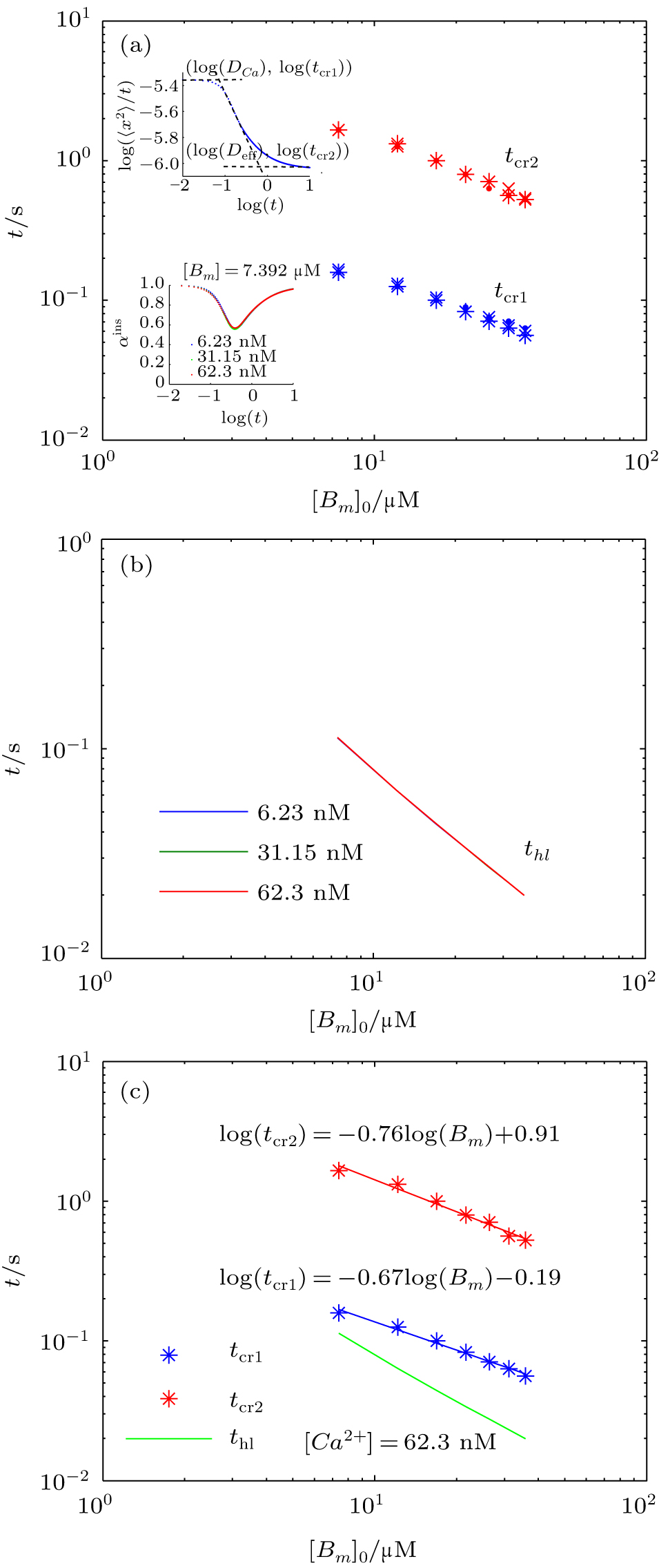 | Fig. 8. (a) The first and second crossover time (tcr1, tcr2) each as a function of mobile buffer concentration at three different concentrations of calcium ions: [Ca2+]0 = 6.23,31.15,62.3 nM (note the overlap of data points for different values of [Ca2+]0), for k+ = 1 and k− = 2.6. The upper inset in panel (a) shows the intersections of the lines yielding the crossover times, tcr1 and tcr2. The lower inset shows αins for different values of [Ca2+]0. (b) The calcium half-life obtained from Eq. ( |
To get an insight into tcr1 and tcr2, we calculate the half-life of calcium concentration in a well-mixed condition where both calcium and buffer are uniformly distributed. For the reversible bimolecular reactions,
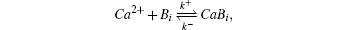
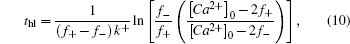
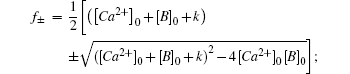
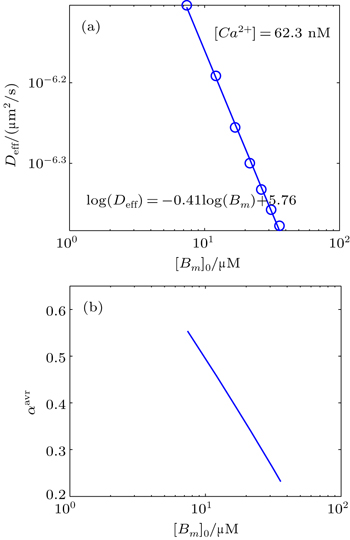
Likewise, we obtain the calcium effective diffusion coefficient Deff (or D(∞)), from the simulation results as plotted in Fig.
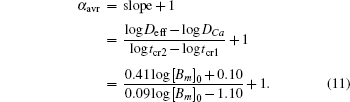
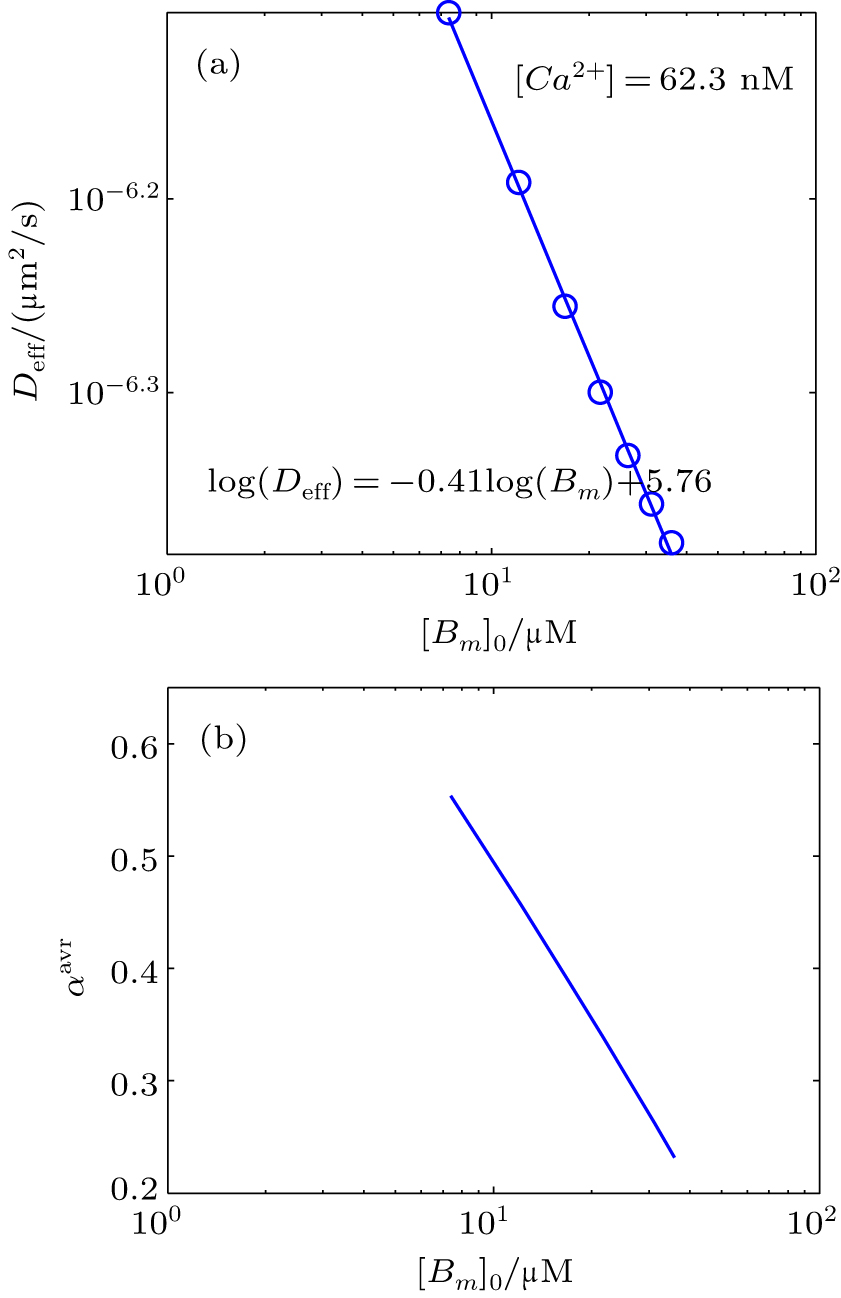 | Fig. 9. (a) Plot of the effective diffusion coefficient (Deff) as a function of initial mobile buffer concentration [Bm]0. (b) The value of αavr in the period of anomalous sub-diffusion calculated from Eq. ( |
In this work, the roles of biochemical reactions in diffusion dynamics are computationally investigated by using both deterministic and stochastic algorithms. Because we cannot track individual particle trajectories in the deterministic simulations, methods to estimate the MSD from deterministic calculations are also investigated. We find that the numerical integration of the PDF best estimates the MSD. Computational and theoretical analyses show that the first-order reaction diffusion dynamics are normal at all times, whereas the second-order reaction diffusion dynamics is more complex and depends on reaction parameters. However, the second-order dynamics is found to follow the anomalous sub-diffusion at least at some times. This work clearly confirms that anomalous diffusion can be a result of only chemical reactions and only second-order reactions can give rise to anomalous diffusion dynamics in a reaction diffusion system.
| 1 | |
| 2 | |
| 3 | |
| 4 | |
| 5 | |
| 6 | |
| 7 | |
| 8 | |
| 9 | |
| 10 | |
| 11 | |
| 12 | |
| 13 | |
| 14 | |
| 15 | |
| 16 | |
| 17 | |
| 18 | |
| 19 | |
| 20 | |
| 21 | |
| 22 | |
| 23 | |
| 24 | |
| 25 | |
| 26 | |
| 27 | |
| 28 | |
| 29 | |
| 30 | |
| 31 | |
| 32 | |
| 33 | |
| 34 | |
| 35 |